Stimulace lymfangiogeneze
51 integrin
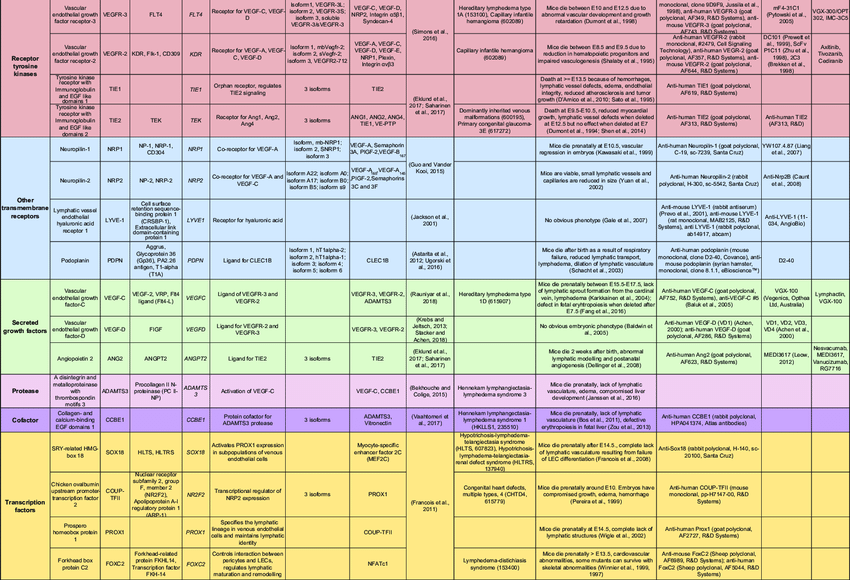
- Co-receptors for VEGFR-3 (Wang et al., 2001; Zhang et al., 2005) [55]
ADAMTS3
Popis
- Originally assumed to be a procollagen II processing enzyme (Fernandes et al., 2001) [41]
Funkce
- VEGF-C cleavage by ADAMTS3 requires the collagen- and calcium-binding EGF domains 1 (CCBE1) protein (Bos et al., 2011; Bui et al., 2016; Jeltsch et al., 2014; Le Guen et al., 2014) [41]
ADAMTS3 mutations
- Can be the cause of hereditary lymphedema conditions (Brouillard et al., 2017; Jha et al., 2017) [41]
Adamts3-deleted mice
- Do not show procollagen processing defects
- Prenatally lethal edema phenotype (Janssen et al., 2015) [41]
AKT 1
Mutace
- Proteus syndrome
- Asi 40 pops. případů
- N/A
- Gain of function [55]
Mice Somatic Akt1–/–
ANG ligands
- Can signal in two different configurations
- TIE receptors from two cells in close proximity can become ligated by ANG ligands (trans complexes)
- TIE receptors can form complexes within one cell
- Triggered by e.g. matrix-bound ANG ligands (cis complexes) (Fukuhara et al., 2008; Saharinen et al., 2008) [41]
ANG2
- Ligand of TIE receptors
- Partial agonist (Yuan et al., 2009)
- Biological response within the ANG/TIE signaling is complex and context-dependent (reviewed by Eklund et al., 2017) [41]
Deletion of Ang2
- Defective collecting lymphatic vessel formation (Dellinger et al., 2008)
- Smaller diameter of lymphatic capillaries
- Without any noticeable effect in the early lymphatic development (Shen et al., 2014)
- Ang1 can rescue the lymphatic abnormalities observed in the Ang2 deleted mice (Dellinger et al., 2008) [41]
ANG2-blocking antibody
- Inhibited embryonic lymphangiogenesis
- Blocked VE-cadherin phosphorylation at tyrosine residue 685
- Concomitant formation of button-like junctions in initial lymphatics
- Defective junctions were associated with impaired lymph uptake [53]
- In collecting lymphatics, adherens junctions were disrupted
- Vessels leaked upon ANG2 blockade or gene deletion
- Suppressed the onset of lymphatic valve formation and subsequent valve maturation [53]
Endothelium-specific ANG2 overexpression
- Induced lymphatic hyperplasia
CCBE1
Popis
Funkce
- Lymphatic development
- CCL21 for dendritic cell migration (Russo et al., 2016; Vaahtomeri et al., 2017a; Weber et al., 2013) [41]
- Enhances the lymphangiogenic effects of VEGFC in vivo [Bos et al., 2011] [4]
- Essential for fetal liver erythropoiesis [Zou et al., 2013]
- Important for lymphatic development
Genetic knock-down in zebrafish
Human mutations
- Lymphangectasia lymphedema syndrome 1 (HKLLS1, 235510) [41]
- Can be responsible for Hennekam Syndrome - human hereditary condition - Hennekam lymphangiectasia lymphedema syndrome 3
- Characterized by generalized lymphedema (Alders et al., 2013, 2009) [41]
CCBE1 homozygous
Compound heterozygous mutations
- Hennekam lymphangiectasia-lymphedema syndrome (OMIM 235510)
- Severe peripheral lymphedema associated with
- Intestinal lymphangiectasias
- Characteristic facial features
- Growth and mental retardation
- Hydrops fetalis [Hennekam et al., 1989; Alders et al., 2009, Alders et al., 2013; Connell et al., 2010] [4]
- All CCBE1-mutated patients clearly had Hennekam syndrome [Alders et al., 2009]
- Future screens of CCBE1 could be restricted to patients with
- Generalized lymphatic dysplasia
- With or without facial anomalies
- With a recessive mode of inheritance [4]
Možnosti ovlivnění
Stimulace CCBE1 u heterozygotů
- ?
Synergie s CCBE1
- Stimulace jeho second messengers / jiných drah
- ?
C-C chemokine ligand 21 - CCL21
Funkce
- CCR7 and its ligand, CCL21
- CCL21/CCR7 signaling is necessary for lymphatic system–dependent cell trafficking from the periphery to
- The organ
- Its corresponding DLNs [42]
Organ injury
- Stimulates enhanced expression of the key C-C chemokine ligand 21 (CCL21)
- LVs provide the route for inflammatory cells expressing the CCL21 receptor CCR7
- Activated dendritic cells (DCs) [42]
- Lymphocytes
Ledviny, novotvorba lymfatických cév a fibroza ledvin
- New injury-induced Lymphatic Veins (LV) expression of CCL21
- Stimulates recruitment of imunitních buněk into RDLNs and spleen
- CCR7 - dendritic cells (DCs)
- Lymphocytes
- Resulting in a systemic lymphocyte expansion
- Injury-induced intrarenal inflammation and fibrosis could be attenuated by
- Blocking the recruitment of CCR7 ? cells into RDLN and spleen
- Inhibiting lymphangiogenesis [42]
- Lymphangiogenesis in the kidney and RDLN (renal draining lymph nodes) is driven by proliferation of preexisting lymphatic endothelium
- Expressing the essential C-C chemokine ligand 21
- Intrarenal lymphangiogenesis
- Occurs at the site of interstitial lesions
- Strongly correlates with fibrosis [42]
- Intrarenal lymphangiogenesis identified in
- Interstitial fibrotic regions in a rat remnant kidney model
- Proteinuria model
- Increased VEGF-C and VEGF-D expression was detected in the kidney [42]
- Potent fibrogenic cytokines via up-regulation of VEGF-C in a unilateral ureteral obstruction (UUO) model including
- Transforming growth factor beta 1
- Connective tissue growth factor (CTGF) [42]
- Blocking VEGF-C/D–VEGFR3 signaling
- Decreased UUO-induced renal lymphangiogenesis [42]
- Following renal injury, lymphangiogenesis occurs
- Via local proliferation of existing endothelium in the kidney and corresponding RDLNs
- Blocking recruitment of CCR7+ cells into RDLNs and the spleen through markedly attenuated intrarenal inflammation and fibrosis
- Inhibition of lymphangiogenesis
- Administration of a CCR7-neutralizing antibody [42]
Plíce
- Important role in directing effector memory T lymphocyte trafficking
- To the lungs via afferent LVs [42]
- Mice lacking CCR7
- Impaired DC and T lymphocyte trafficking
- Fail to mount an adaptive immune response [42]
Peripheral solid organ lymphangiogenesis
- In association with various inflammatory conditions including
- Transplant rejection,
- Hypertension,
- Matrix stiffness,
- Myocardial infarction,
- Tumor metastasis [42]
Kůže
- CCR7 was initially reported to be necessary for T cell entry into lymphatics from skin [51]
CLEC2
COUP-TFII
- Chicken ovalbumin upstream promoter-transcription factor 2
- Nuclear receptor subfamily 2, group F, member 2 (NR2F2), Apolipoprotein A-I regulatory protein 1 (ARP-1) [41]
- Transcriptional regulator of NRP2 expression
- 3 isoforms
- Major interacting protein: PROX1 [41]
Mutace genu NR2F2
- Congenital heart defects, multiple types, 4 (CHTD4, 615779) [41]
Mice mutace
- Mice die prenatally around E10. Embryos have compromised growth, edema, hemorrhage (Pereira et al., 1999) [41]
Calcium-binding EGF domains 1
Cxcl12a
FGF-2 - Basic fibroblast growth factor–2
- Promote lymphangiogenesis
- By interacting with the LV endothelial hyaluronan receptor 1 (LYVE-1) transmembrane domain
- A process that is independent of the activation of VEGFR3 signaling
www.researchgate.net/publication/334060542_Lymphangiogenesis_in_kidney_and_lymph_node_mediates_renal_inflammation_and_fibrosis
FGFR-3
Funkce
- LECs showed enhanced cell proliferation [48]
- Signifficant contribution of FGFR3 to the flow-activated LEC proliferation [48]
Knockdown of FGFR3
- Reduced the flow-activated LEC growth
Upregulated FGFR3
- May make the cells more sensitive to its limited ligands, such as FGF2 [48]
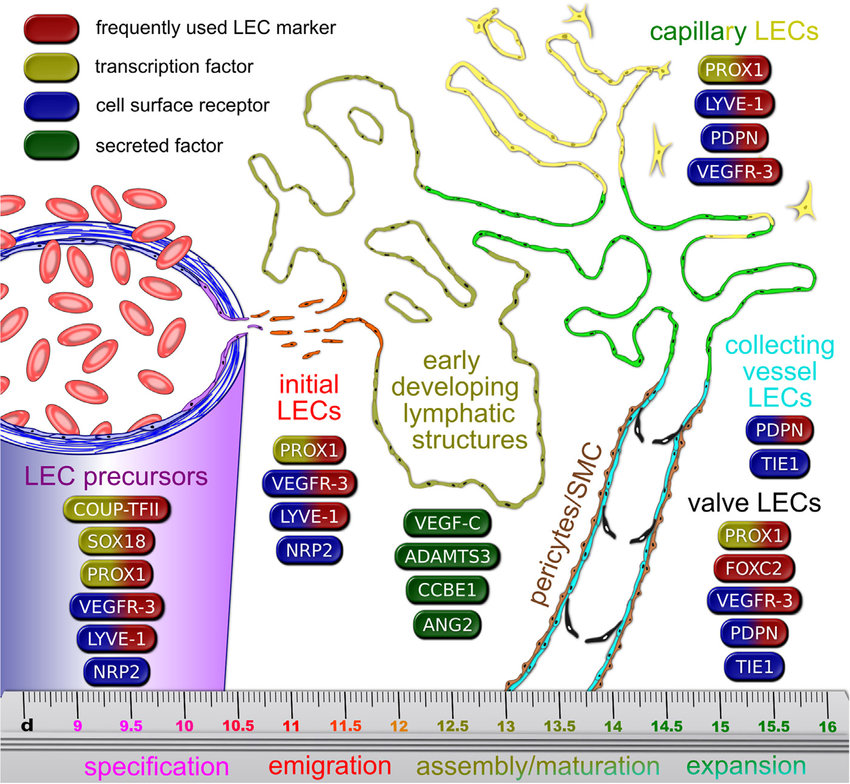
FOXC2
- Forkhead box C2
- Transcription factor
Funkce
- FOXC2 suppresses PDGFB production [Shimoda et al., 2011]
- Controls the interaction between pericytes and LECs (Petrova et al., 2004)
- Becomes important during later stages of the lymphatic development.
- Required together with NFATc1 for the lymphatic remodeling and maturation
- Formation of lymphatic valves in the precollectors and collectors (Norrmén et al., 2009) [41]
- Regulates the expression of genes involved in
- Cell growth
- Proliferation
- Differentiation
- Longevity [Fang et al., 2000]
Mutace FOXC2
- In puberty or late-onset primary lymphedema associated with distichiasis (LDS, OMIM 153400) [4]
- Hereditary lymphedema II (OMIM 153200) [5]
- Often associated with
- Distichiasis (double row of eyelashes)
- High penetrance
- Not all patients with this feature carry a mutation in FOXC2
- Sometimes ptosis (OMIM 153400)
- Lymphedema-distichiasis syndrome (153400) [41]
- And/or yellow nails (OMIM 153300) [5]
Foxc2–/– mice
- Abnormal lymphatic patterning
- Arrested lymphatic valve development [5]
Foxc2–/+ mice - heterozygotes
- increased recruitment of pericytes hampers the function of collecting lymphatics
- Reminiscent of the human phenotype [5]
Genetika
- Majority of the FOXC2 mutations are
- Insertions,
- Deletions
- Nonsense mutations
- Leading to
- MRNA decay
- Truncated loss-of-function proteins [Dagenais et al., 2004; Ghalamkarpour et al., 2009a; van Steensel et al., 2009] [4]
- FOXC2 suppresses PDGFB production [Shimoda et al., 2011]
Loss of FOXO2 activity
- accumulation of vascular smooth muscle cells in collecting lymphatics of knock-out mice and also in patients [Petrova et al., 2004; Norrmen et al., 2009] [4]
- Cca 7% lymfedémů [1]
Diagnostické možnosti v praxi
- ?
Potencionální terapie
Snížení produkce PDGFB v lymfat. cévách
- ?
Potlačení množení smooth muscle cells /pericites in collecting lymphatics
- ?
GATA2 - GATA-binding protein 2
- Zinc finger transcription factor
Funkce
- Putative enhancer element upstream of the key lymphatic transcriptional regulator PROX1
- That is bound by GATA2, and the transcription factors FOXC2 and NFATC1
- Cross-regulation
- Transcription factors modify expression levels of several other proteins
- Some involved in lymphangiogenesis
- Transcription factor that controls PROX1 and FOXC2 expression
- PROX1 regulates FLT4, and FOXC2 controls proteins essential for lymphatic valves, such as connexins [5]
- Crucial for hematopoietic stem cell development during both embryogenesis and adulthood
- In hematopoietic progenitor cells
- Key roles during lymphovenous and lymphatic vessel valve formation
- Regulate the transcription of genes that are important for lymphatic vessel valve development
- Important for vascular integrity and separation of the blood and lymphatic vascular networks [34]
- GATA2 transcription factor
- Critical regulator of matrix stiffness
- Induced transcriptional program in the LECs
- Activation of GATA2 by increased mechanical stimulus
- LECs to stiff matrix or oscillatory flow
- GATA2 is required for LEC emigration from the veins
- LECs assemble in the jugular region of the embryo into the first primitive vessels
- The primordial thoracic ducts (pTD)
- Peripheral longitudinal lymphatic vessels (PLLV)
- From which most peripheral lymphatic vascular beds form by vessel sprouting [39]
- GATA2 regulates a stiffness-induced transcriptional program
- GATA2 controls LEC responsiveness to VEGF-C
- Respond to ECM-derived mechanical signals in blood ECs (BECs) by changing its nuclear/cytoplasmic localization [47]
Heterozygous germline mutations
- Range of clinical phenotypes
- Catalogue of human GATA2 mutations results in lymphedema
- Loss of Gata2 does not prevent LEC differentiation but inhibits migration of venous-derived LEC progenitors
Emberger GATA2 missense mutants - Emberger syndrome
- Result in complete loss of GATA2 function (key to lymphedema developement)
- Profoundly reduced capacity to bind this element - PROX 1
- Identified in patients with primary lymphedema with myelodysplasia - Emberger syndrome (OMIM 614038) [Hahn et al., 2011; Ostergaard et al., 2011b] [4]
- Lymphedema and predisposition to
- Myelodysplastic syndrome (MDS, OMIM 614286)
- Acute myeloid leukemia (AML, OMIM 601626)
- Monocytopenia with mycobacterial infection syndrome (MonoMAC, OMIM 614172)
- Mostly to: dendritic cell, monocyte, B lymphocyte and natural killer lymphocyte deficiency (DCML) [Kazenwadel et al., 2012] [4]
- Japanese patient with a GATA2 mutation
- MonoMAC and Emberger syndromes [Ishida et al., 2012] [4]
- no obvious genotype-to-phenotype correlations [Hyde and Liu, 2011; Holme et al., 2012] [4]
- Suggesting that modifiers play a role [4]
Most GATA2 missense mutations incl. prevalent T354M
- Do not correlate with lymphedema
- Hypothesis that complete heterozygous loss of GATA2 function underlies lymphedema was complicated by the description of Emberger syndrome patients with missense mutations
- R361L, C272R , R396Q [34]
Conditional Gata2 deletion in mice
- GATA2 is required for both development and maintenance of lymphovenous and lymphatic vessel valves
SNPs in GATA2
- Associated with
- Coronary artery disease
- In arterial development
Ablation of gata2 in zebrafish
- Affects morphogenesis of the dorsal aorta
Gata2 “null” haploinsufficiency
- Propensity to develop lymphedema
- Through regulation of genes, including Prox1 and Foxc2
- That are important for lymphatic vessel development and valve development
siRNA-mediated GATA2 knock-down in primary embryonic mouse LECs
- Reduced PROX1 levels and FOXC2 [34]
Adult heterozygous Gata2 LEC/+ mice
- Injected with Evans Blue dye
- Collecting lymphatic vessels of substantially larger caliber than controls
- Reduced transport of Evans Blue dye to the thoracic duct
- Blood within the thoracic duct
Možnosti Stimulace GATA2 za účelem zmírnění lymfedému
NOTCH1/RBJ-kappa
- Required to initiate Gata2 expression in hematopoietic stem cells in the embryonic aorta-gonad-mesonephros region
Shear stress - tření, tok lymfy
Takže manuální lymfodrenáž a lymfotaping, bandáže
BMP signalling
- Required to induce Gata2
- Shown to control lymphatic vessel valve formation
Měkká matrix
- GATA2 expression is increased in LECs grown on a soft matrix
- GATA2 is required for early lymphatic vascular morphogenesis
- By controlling lymphangiogenic growth factor responsiveness
- Following the exposure of migrating venous-derived LECs to a soft embryonic tissue [39]
Inhibice GATA2 - zhoršení lymfedému
Retinoic acid signalling (Vitamín A !!!)
- Impact on the transcriptional activity of GATA2
- Direct interaction between the zinc fingers of GATA2 and the DNA-binding domain of RA receptor alpha (RAR alpha)
- Aberrant development of LVVs in Cyp26b1–/– mice
- RA signalling is elevated
Mutagenní účinek vysokých hladin vit. A ???
Loss of NOTCH1 results in
- Fewer valves
- Disrupted reorientation of valve endothelial cells
- Reduced levels of valve markers, including
- ITGalpha9
- FN-EIIIA
Notch-induced gene Hes1
- Negatively regulates Gata2 in hematopoietic stem cells of the AGM
- Controlling the production of functional HSC
- NOTCH1 function has recently been shown to be important for lymphatic vessel valve development
GATA1
- Repress GATA2 expression in hematopoietic cells
- GATA1 is both positively and negatively regulated by the Notch signalling pathway [34]
Stimulace PROX1, FOXC2, FLT4
- ?
GJA1 (encoding connexin-43)
Funkce
- Highly enriched on the upstream side of lymphatic valves
- Forms hemi- and intercellular [5]
Mutace - Amino acid substitution
- Lymphedema, as part of the oculodentodigital dysplasia (OMIM 164200)
- Oculodentodigital dysplasia/lymphedema
- Affects the eyes, face, teeth, and digits [5]
- V.s. specifically alters channel properties leading to valve dysfunction [5]
Homozygous inactivation of Cx43 in mice
- Is lethal at birth
- Cardiac malformation [5]
GJA1 (encoding connexin-43)
Funkce
- Highly enriched on the upstream side of lymphatic valves
- Forms hemi- and intercellular [5]
Mutace - Amino acid substitution
- Lymphedema, as part of the oculodentodigital dysplasia (OMIM 164200)
- Affects the eyes, face, teeth, and digits [5]
- V.s. specifically alters channel properties leading to valve dysfunction [5]
Homozygous inactivation of Cx43 in mice
- Is lethal at birth
- Cardiac malformation [5]
GJC2 (encoding connexin-47)
Funkce
- CX47 is expressed in
- Lymphatic endothelial cells
- On the upstream side of lymphatic valves
Nonsynonymous mutations in GJC2
- Discovered in a few families with
- Late-onset (age 30) autosomal dominant lymphedema (OMIM 613480)
- All 4 extremities (‘four-limb lymphedema’)
- Sometimes associated with
- Saphenous vein insufficiency
- Blepharoptosis
- Involvement of the face or genitalia
- Recurrent cellulitis
- Some families showed reduced penetrance [Ferrell et al., 2010; Ostergaard et al., 2011a] [4]
GJC2 amino acid substitutions
- Alter but do not abolish connexin function [5]
- Likely cause gain-of-function [4,5]
- Substitutions of highly conserved amino acids in connexin 47 (CX47) cause
- Lymphedema in all four extremities
Cx47 homozygous knockouts
- Have no lymphatic defect [5]
Loss-of-function mutations in GJC2
- Found in patients with inherited autosomal recessive Pelizaeus-Merzbacher-like disease (PMLD, OMIM 608804)
- Primarily premature stop codons
- Hypomyelinating disorder of the central nervous system [Uhlenberg et al., 2004] [4]
- Hypomyelinating leukodystrophy 2 (OMIM 608804) without lymphedema [5]
GJC2 (encoding connexin-47)
Funkce
- CX47 is expressed in
- Lymphatic endothelial cells
- On the upstream side of lymphatic valves
Nonsynonymous mutations in GJC2
- Discovered in a few families with
- Late-onset (age 30) autosomal dominant lymphedema (OMIM 613480)
- All 4 extremities (‘four-limb lymphedema’)
- Sometimes associated with
- Saphenous vein insufficiency
- Blepharoptosis
- Involvement of the face or genitalia
- Recurrent cellulitis
- Some families showed reduced penetrance [Ferrell et al., 2010; Ostergaard et al., 2011a] [4]
GJC2 amino acid substitutions
- Alter but do not abolish connexin function [5]
- Likely cause gain-of-function [4,5]
- Substitutions of highly conserved amino acids in connexin 47 (CX47) cause
- Lymphedema in all four extremities
Cx47 homozygous knockouts
- Have no lymphatic defect [5]
Loss-of-function mutations in GJC2
- Found in patients with inherited autosomal recessive Pelizaeus-Merzbacher-like disease (PMLD, OMIM 608804)
- Primarily premature stop codons
- Hypomyelinating disorder of the central nervous system [Uhlenberg et al., 2004] [4]
- Hypomyelinating leukodystrophy 2 (OMIM 608804) without lymphedema [5]
Hepatocyte growth factor - HGF
- Independently identified for its potent effects
- On cell motility in epithelial cells (scatter factor )
- Cell proliferation in hepatocytes (hepatocyte growth factor/hepatopoietin A22–24) [1]
- Large multidomain polypeptide
- Similarities to plasminogen
- Synthesized as a single long polypeptide
- Requires proteolytic conversion to its active form as a heterodimer
- Biologic activity is dependent on its binding to the MET receptor [1]
- Lymphatic endothelial cells (LEC) expressed the receptor MET for HGF
- HGF/MET signaling affects
- Morphogenic differentiation
- Cell motility
- Growth
- Intercellular junctions
- Survival [1]
Mutace HGF
- In primary lymphedema
- Lymphedema/lymphangiectasia [55]
- Breast cancer-associated secondary lymphedema
- Pathway provides a new target for the prevention and/or treatment of lymphedema [1]
- Confers susceptibility to secondary lymphedema and other syndromic lymphatic variation [1]
HGF terapie
- Mouse corneal lymphangiogenesis model
- HGF could be lymphangiogenic [1]
Treatment of LEC with recombinant HGF promoted
- Proliferation
- Migration
- Tube formation of LEC [1]
Subcutaneous or transgenic delivery of HGF
- Promoted lymphatic vessel formation in the mouse !!
- HGF/MET as viable candidate genes for hereditary lymphedema [1]
HGF Injection of a plasmid expressing HGF
- In rat tail
- In a mouse model of upper limb lymphedema that simulates breast cancer-related lymphedema
- Prevented overt development of lymphedema
- Stimulated development of new lymphatic vessels in the long term
- Expression of HGF by plasmid transfer
- Promotes lymphangiogenesis
- Ameliorates secondary lymphedema in the rat tail injury model [1]
HRAS
Mutace
- Costello syndrome/chylous ascites
- Few with chylous ascites
- Low penetrance
- Gain of function
AD Hras–/–
IKBKG (NEMO)
Funkce
- An NF-kappa B modulator
- IKBKG to activate NF-kappa B
- NF-kappa B upregulates PROX1
- Cooperates with it to induce VEGFR-3 expression [5]
- Modulate the activity of transcription factors
- Affect nuclear dynamics
Mutations in patients
- Complex lymphedema syndromes
- Rare X-linked syndrome
- Anhydrotic ectodermal dysplasia with
- Immunodeficiency,
- Osteopetrosis
- Lymphedema (OLEDAID; OMIM 300301) [5]
- Associated with
- Incontinentia pigmenti in the mother (OMIM 308300)
- Five cases of OLEDAID with an IKBKG mutation have been reported
- Loss-of-function mutations
- Cause incontinentia pigmenti
- OLEDAID-causing mutations are hypomorphs
- Diminish but do not abolish the ability of IKBKG to activate NF-kappa B
Males and homozygous Ikbkg–/– murine females
- Embryonic lethal in
- Die from severe apoptosis
ITGA9
Mutace
- Fetal chylothorax
- Popsaných případů 6
- Penetrance High
- Missense [55]
AR, de novo Itga9–/–
Intracellular calcium dynamics
- In cultured LECs
- Depends on the magnitude of the shear stress
- Blockage of calcium release–activated calcium channels
- Signi?cantly reduced the calcium mobilization [47]
KIF11 gen - kóduje EG5 protein
Funkce
- EG5 acts as a homotetrameric kinesin motor
- Kinesin family member 11, a DNA-interacting protein
- Members of this protein family are involved in
- Establishing a bipolar spindle during mitosis for chromosome positioning and centrosome separation [5]
- Mitotic spindle assembly and function [Ostergaard et al., 2012] [4]
- inhibition of EG5 activates the PI3K/AKT pathway [5]
KIF11 mutations
- MCLMR (microcephaly with or without chorioretinopathy, lymphedema, or mental retardation, OMIM 152950) [Ostergaard et al., 2012] [4]
- Mutations are predicted to result in loss-of-function of EG5 [5]
- Can be sporadic or inherited as an autosomal dominant trait
- MLCRD (microcephaly, lymphedema, chorioretinal dysplasia)
- CDMMR (chorioretinal dysplasia, microcephaly and mental retardation)
Heterozygous KIF11 mutations
- Lower limb lymphedema of variable expressivity associated with:
- Microcephaly with or without chorioretinopathy
- Mental retardation (MCLMR; OMIM 152950) [5]
Kif11+/– mice
- Phenotypically normal [5]
Kif11–/– mice
- Die prior to implantation [5]
KLF2/4
- Induces proliferation [47]
- Both KLF2 and KLF4
- Bind to the 210-kb upstream area of the VEGF-C gene
- Bindings were profoundly increased by laminar flow [47]
- Loss of epigenetic KLF4-mediated transcriptional suppression
- Crucial for upregulation of VEGF-A in breast cancer cells [55]
KLF2
- Inhibits VEGF-mediated angiogenesis
- Laminar shear stress inhibits endothelial cell metabolism via KLF2-mediated repression of PFKFB3
- Induces a gene expression pattern that can be seen in functionally quiescent endothelial cells
- Suppresses angiogenesis of liver endothelial cells through ERK1/2
- Decreased the hypoxia-induced VEGF protein level in HUVECs
- Activate VEGF/VEGFR2 signaling and survival of HUVECs in response to laminar ?ow
- Cooperates with a ETS family protein ERG to activate Flk1/VEGFR2 expression during vascular development [47]
Knockdown of KLF2 and KLF4
- Caused largely additive effects on the ?ow-induced regulation of the genes [47]
- Single knockdown of each gene did not clearly alter the ?ow-induced activation of cell cycle progression of LECs or BECs
Adenoviral overexpression of KLF2 or KLF4 in LECs
- Significantly upregulated VEGF-A, VEGF-C, and FGFR3
- While downregulating p57 [47]
KRAS
Mutace
- Noonan syndrome, cardiofaciocutaneous
- Syndrome/chylothorax
- Few with chylothorax
- Low penetrance
- Gain of function [55]
AD Kras–/–
LYVE-1 n
- Cell surface glycoproteins LYVE-1 and podoplanin (PDPN)
- Good indicators of lymphatic nature [41]
Exprese LYVE-1
- Liver BECs (Carreira et al., 2001)
- Certain macrophages (Schledzewski et al., 2006)
- Generally a useful marker to identify lymphatic capillaries (Banerji et al., 1999)
- Decreased on lymphatic pre-collectors
- Absent from collectors (Lutter et al., 2012) [41]
Funkce
- Receptor for hyaluronic acid
- Functions in dendritic cell entry into the lymphatics (Johnson et al., 2017) [41]
- Internalizes hyaluronic acid (HA)
- HA is soluble within the extracellular matrix
- Found in lymph
- Lymph node participates in its turnover
- Quite high
- Up to one-third of the total HA pool turning over each day [51]
- Functional role for LYVE-1 on the endothelium remained elusive [51]
- HA component of the cell wall of group A streptococci
- Observed to bind ef?ciently to LYVE-1 in lymphatic endothelial cells
- Binding mediated dissemination of the microorganism to lymph nodes
- Clustering or multimerization of HA on the surface of microorganisms or cells
- Allows for LYVE-1 binding on lymphatic endothelium
- Interacting with interstitium-derived HA during its degradation in lymph and lymph nodes
- LYVE-1 may serve as a microbial recognition receptor.
- LYVE-1 on lymphatic endothelial cells is involved in adhesion to cells that assemble multimeric HA on their surfaces such as
- DCs
- Macrophages [51]
- Subpopulation of macrophages that express LYVE-1 as well and exhibit an alternatively activated phenotype that is proangiogenic [51]
Fragments of HA
- Trigger DC migration from the skin
- Associated with in?ammation and tissue injury
- Implicated in the excess accumulation of neutrophils and other immune cells in various clinical contexts
- Transplant rejection [51]
Záněty
- Expansion of lymphatic capillaries, through lymphangiogenesis
- Observed at sites of in?ammation in many models and in?amed tissues
- Transplants [51]
- Lymphangiogenesis in the transplanted lung
- Protects against rejection by promoting clearance of HA from the grafted organ [51]
Laminar flow in the vessels
- Has been shown to promote vessel sprouting through inhibition of NOTCH1 [47]
- Laminar flow downregulates Notch activity to promote lymphatic sprouting
- Upregulation of KLF2, KLF4, and eNOS, in response to the low-rate laminar ?ow condition [47]
- Higher rate laminar ?ows, which are comparable to blood ?ow, instead suppress proliferation of vascular endothelial cells [47]
Locus in 15q
Mutace
- Cholestasis-lymphedema syndrome
- Aagenaes syndrome
- AR – [55]
Multipotent adult progenitor cells (MAPCs)
- Potential to diferentiate down the lymphatic endothelial lineage
- Trophically supported lymphatic endothelial cell behaviour in vitro
- In vivo, MAPC transplantation supported
- Blood vessel and lymphatic capillary growth in wounds
- Restored lymph drainage across skin flaps
- By stimulating capillary and pre-collector vessel regeneration
- Mediated survival and functional reconnection of transplanted lymph nodes to the host lymphatic network by
- Improving their (lymph) vascular supply
- Restoring collector vessels
Stimulace MAPCs
- Upon VEGF-A exposure human (h)MAPCs
- Differentiate into arterial and venous endothelial cells
- Gain general endothelial cell marker expression
- Prospero homeobox 1 (Prox1), the lymphatic master switch, was signifcantly induced
- Expression of additional lymphatic genes (i.e., Pdpn and Itg9a) is upregulated by forced Prox1 expression
- A fraction (21 ± 6%) of VEGF-A-exposed MAPCs also expressed
- Lymphatic Vessel Endothelial Hyaluronan Receptor 1 (LYVE1)
- Culturing MAPCs with VEGF-A results in a mixture of arterial, venous or lymphatic endothelial cells
- In ischaemic limbs
- MAPCs had a limited direct contribution to blood vascular endothelium
- 72 hour mMAPC- or hMAPC-conditioned media signifcantly stimulated
- Lymphatic endothelial cell sprouting
- Proliferation
- Migration
- VEGF-A
- Angiopoietin-2 (ANG-2)
- VEGF-C expression
- MMAPCs (mouse) and hMAPCs (human) had a 62% overlap in their cytokine/growth factor secretion
- HMAPCs
- Secreted larger amounts
- Broader complement of these factors [39]
- HMAPCs applied onto circular wounds
- Accelerated wound closure
- Homogenous engrafment in the wound bed
- Only occasional in situ diferentiation to (lymphatic) endothelial cells [39]
MAPC transplantation
- Represents a promising remedy for lymphatic system restoration at different anatomical levels
- Treatment for lymphedema
- Wound healing [39]
- Transplanted MAPCs, rather than being incorporated into lymphatic vessels
- Strategically positioned themselves
- Near lymphatic vessels
- Near transplanted lymph nodes
- From where they could deliver a complement of trophic factors supporting lymphatic regeneration [39]
- Some persisting up to 6 months
- HMAPCs secrete significant levels of VEGF-A
- Trophic efects of mesenchymal stem cells on lymphatic endothelial cells
- MAPCs support regeneration of the lymphatic tree at di?erent anatomical levels
- Many other candidates may contribute
- Angiopoietins,
- Hepatocyte growth factor
- Insulin growth factor binding proteins
- (tissue inhibitors of ) matrix metalloproteinases
- Stromal cell-derived factor-1
- Interleukin-8
- Osteopontin,
- Amphiregulin [39]
- Significantly accelerated wound
- Re-epithelialisation,
- Granulation tissue formation
- Collagen remodelling [39]
MET receptor for HGF
- Predominantly in cells of mesenchymal and epithelial origin [1]
- Prototypical receptor tyrosine kinase
- First recognized as a protein product of an oncogene, TPR-MET
- MET signals are channeled by an unconventional multi-docking site
- That consists of two tyrosines that
- When phosphorylated, recruit a wide spectrum of transducers and adaptors
- PI3K, Src, Grb2, Shp2 Gab1, and STAT3
- Whether mutations in any of these downstream genes acting in the HGF/MET signaling pathway are responsible for lymphatic phenotypes has not been studied.
- Extracellular Sema domain
- Necessary for MET receptor dimerization and activation in vitro using several human cell lines [1]
MET receptorové mutace a klinika
- Rs3458946
- Lymphedema and protein losing enteropathy secondary to intestinal lymphangiectasia [1]
Motif 3/collagen
NFATC1
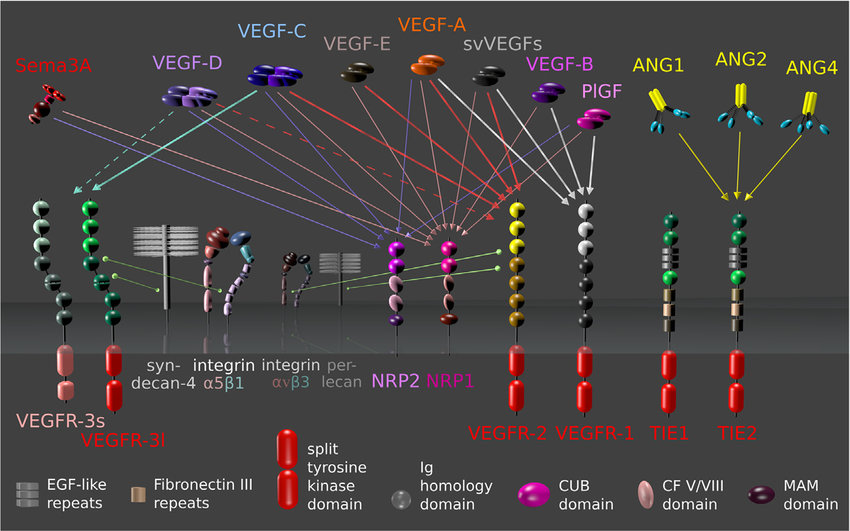
Neuropilin-1 (NRP1) and neuropilin-2 (NRP2)
- First described as transmembrane proteins of neuronal cells
- Regulate the growth of dendrites and axons together with their different semaphorin ligands
- Attractants or repellents (Schwarz and Ruhrberg, 2010)
- Expressed by endothelial cells
- NRP1 more prominently on arteries
- NRP2 more prominently on LECs and veins (Herzog et al., 2001; Yuan et al., 2002)
- Act as co-receptors for VEGF ligands
- Stabilizing the growth factor/receptor complex
- Likely do not exercise a signaling function in endothelial cells (Guo and Vander Kooi, 2015)
- All VEGF family members have been seen to interact with NRP1 (and most with NRP2)
- VEGF-A/NRP1 (Kawamura et al., 2008)
- VEGF-C/NRP2 (Xu et al., 2010)
- Appear to be significant both in vivo and in vitro (Karpanen et al., 2006)
ORAI1
- A pore subunit of the calcium release-activated calcium (CRAC) channel
- Identified to induce the shear stress phenotypes and cell proliferation in LECs responding to the fluid flow
- ORAI1 induced upregulation of KLF2 and KLF4 in the flow-activated LECs
- Low-rate steady laminar ?ow activates a highly selective calcium channel ORAI1 to upregu-late Krüppel-like factors (KLF) 2 and 4, which directly regulate the genes promoting cell proliferation and survival [48]
- Two KLF proteins cooperate to regulate VEGF-A, VEGF-C, FGFR3 and p57
- By binding to the regulatory regions of the genes [48]
- Important calcium release–activated calcium channel responsible for the store-operated Ca2+ entry process [48]
Freshly isolated LECs from Orai1 knockout embryos
- Reduced expression of KLF2, KLF4, VEGF-A, VEGF-C, and FGFR3
- Elevated expression of p57
Mouse embryos deficient of Orai1, Klf2, or Klf4
- Significantly reduced lymphatic density
- Impaired lymphatic development [48]
ORAI1 knockdown in LECs
- Strongly reduced the ?ow-induced expression of VEGF-C and FGFR3
- Marginal suppression of VEGF-A only at 12 hours [48]
- Reduced expression of VEGF-A, VEGF-C, KLF2, KLF4, and FGFR3, as well as upregulation of p57, in Orai1 KO LECs [48]
- Orai1 KO embryos displayed signi?cant defects in lymphatic development with reduced numbers of LECs [48]
ORAI1 inhibition in LECs
- Abolished the ?ow-induced regulation of VEGF-A, VEGF-C, FGFR3, and p57
- Ef?ciently reversed the cell proliferation activated by laminar ?ow [48]
PECAM-1
- Expressed in lymphatic endothelial cells
- PECAM-1 may contribute to maintenance of vascular endothelial integrity in disease
- Association of PECAM-1 with openings between intercellular junctions instead of with the junctions themselves [47]
PECAM-1 – null mice
- Viable
- Normal vasculature
- Reduced transmigration of leukocytes
- Increased bleeding times and vascular leakage
- Greater susceptibility to endotoxic shock [47]
- PECAM-1 blockade or deletion predisposes FVB/n mice to chronic pulmonary inflammation and fibrosis [47]
- Loss of PECAM-1 results in impaired leukocyte e? ux from blood vessels [47]
C57BL/6 PECAM-1 – null mice
- no change in the distribution of VE-cadherin at buttons or in the integrity of buttons in initial lymphatics [47]
PI3K/AKT pathways
Mutace
- CLOVES,
- Klipple-Trenaunay-Weber syndrome
- Cca 3 případy
- N/A
- Gain of function [55]
Mice - Somatic palpha100-KI
PIEZO1
- Dispensable for early events in valve development, as the up-regulation of FOXC2 and NFATc1 [47]
PTEN
Mutace
- Proteus syndrome,
- Pten hamartoma
- Kolem 10-ti případů
- Penetrance Medium
- Tumor syndrome
- Loss of function
AD, de novo (Pten–/–)
PTN11 - SHP2
Mutace
- Gain of function
- Nad 100 případů popsáno
- Pentrance Medium
- Noonan syndrome 1 (54% with lymphedema) [55]
AD Shp2–/– myši
PTPN14
Funkce
- An intracellular tyrosine-phosphatase [55]
- Recruited to the VEGFR-3 receptor upon VEGF-C stimulation
- Co-immunoprecipitation upon activation by VEGFC [4]
- Dynamic subcellular localization in vitro
- Nuclear in proliferating cells [4]
- Concentrated at intercellular junctions in confluent cells [Wadham et al., 2000; Benzinou et al., 2012] [4]
- Overlap in expression of PTPN14 and VEGFC
- VEGFC stimulation enhances recruitment of PTPN14 to a complex including VEGFR3 [4]
PTPN14 mutations
- Probably explain only a very small proportion of primary hereditary lymphedema [4]
Intragenic deletion encompassing both sides of exon 7 of PTPN14
- Causes a frameshift p.Ser194Argfs*19
- Identified in a consanguineous family with
- Autosomal recessive choanal atresia and
- Lymphedema (OMIM 613611) [Au et al., 2010] [4]
Homozygous deletion of exon 7
- Shift in the reading frame and the appearance of a premature termination codon
- Probably leading to nonsense-mediated mRNA decay and loss of function
- In a single family with juvenile-onset lymphedema and
- Choanal atresia (OMIM 608911)
- Developmental delay
- Pericardial effusion in some affected individuals [Au et al.,2010] [4]
Ptpn14-deficient mice
- Mimic the human phenotype
- Develop lymphedema postnatally due to hyperplastic vessels
- Hyperactive VEGFR-3 signaling due to loss of the phosphatase také narušuje normal lymphatic development [55]
Možnosti ovlivnění
Synergie s PTPN14
- ?
Stimulace PTPN14
- ?
Inhibice blokátorů PTPN14
- ?
Inhibitory VEGFR-3 signalizace
- ?
Perlecan
- VEGFR-2 signalling can be enhanced by interaction (Zoeller et al., 2009).
Podoplanin - PDPN
Exprese
- Immunohistochemical detection of lymphatics (Breiteneder-Geleff et al., 1999) [41]
- Osteoblastic lineage cells (Wetterwald et al., 1996)
- Podocytes
- Important for the formation of the glomerular filtration barrier of the kidney (Matsui et al., 1999)
- Not required for the early steps of lymphatic development
- Absent from the first LECs that emigrate from the cardinal veins (initial LECs or iLECs) (Hagerling et al., 2013) [41]
Prox1 stimulace
- Transcripční faktor
- COUP-TFII and PROX1 continue to be strongly expressed in established lymphatic vessels (Francois et al., 2011) [41]
- Only PROX1, but not COUP-TFII is necessary for the maintenance of the lymphatic identity (Johnson et al., 2008; Lin et al., 2010) [41]
- PROX1 is expressed by several cell types
- Liver cells
- Many stem cells
- In the vascular compartment specific for lymphatic endothelial cells (Bazigou et al., 2011)
- Cardiac (Rodriguez-Niedenführ et al., 2001) valves [41]
- Key transcription factor for the early steps of LEC differentiation from the embryonic veins (Wigle and Oliver, 1999) [41]
- Remains required for lymphatic identity (Johnson et al., 2008) [41]
- Over-expression of PROX1 in BECs modifies their expression patterns to resemble LECs (Hong et al., 2002; Petrova et al., 2002) [41]
- Upregulation of the gene encoding VEGFR-3 [41]
- Seen in the cardinal vein endothelial cells which are committed to LEC differentiation (Wigle et al., 2002) [41]
RAF1
Mutace
- Noonan syndrome 1/lymphangiectasia
- Few with lymphangiectasia
- Low penetrance
- Gain of function [55]
AD Raf1-KI
RAS/MAPK pathways
Inhibitory RAS/MAPK pathway
- Activated in some forms of syndromic lymphedema (Noonan syndrome) and CM-AVM syndrome
- Many inhibitors of this pathway are being developed in laboratories and tested in preclinical trials for cancer therapy
- Some of them may be applicable as therapies for syndromic lymphedema [55]
- Germline or somatic mutations activating the PI3K/AKT signaling pathway
- Small-molecule inhibitors may become useful as treatment
- Many such inhibitors exist
- Some are in clinical use for cancer, including the
- MTOR inhibitor rapamycin
- Preliminary results on PTEN lesions and lymphatic anomalies are encouraging [55]
RASA1
Mutace
- CM-AVM/lymphedema
- Few with lymphedema
- Nízká penetrance
- Loss of function [55]
Myši AD (Rasa1–/–)D
Recombinant VEGF-C
Animal models of secondary lymphedema Szuba and colleagues
- Single 100 ug dose of recombinant human VEGF-C
- Into the surgical bed of a rabbit ear model of lymphedema
- Resulted in histologic evidence of
- Lymphangiogenesis,
- Decreased dermal hypercellularity,
- Improvements in Indocyanine Green (ICG)-based quantifications of lymphatic function
(Szuba et al., 2002)
Follow-up studies mouse tail model of lymphedema
- Recombinant human VEGF-C improved outcomes by increasing lymphangiogenesis
- Improving the morphology of cutaneous lymphatic vessels,
- Ability of lymphatic channels to transport immune cells. (Cheung et al., 2006; Jin et al., 2009)
- www.frontiersin.org/articles/10.3389/fphar.2022.828513/full
Topical formulations of VEGF-C
- To avoid direct injections of the growth factor and to elicit sustained release
- Acidic gelatin hydrogels
- To generate a sustained release of biologically active VEGF-C. (Hwang et al., 2011)
- Combining VEGF-C hydrogel with extracorporeal shock wave therapy or adipose-derived stem cells (ADSCs)
- Even more effective in reducing edema and increasing lymphangiogenesis. (Kim et al., 2013)
- www.frontiersin.org/articles/10.3389/fphar.2022.828513/full
- Recombinant VEGF-C is expensive
Gene therapy methods to increase VEGF-C expression in lymphedema models
Local delivery of naked plasmid VEGF-C DNA in the rabbit ear or mouse tail models
- Increases VEGFR3 expression
- Decreases swelling and fibroadipose tissue deposition
- Improves lymphatic function as assessed using lymphoscintigraphy. (Yoon et al., 2003)
- Similar results were noted by Liu et al. using a rat hindlimb model of secondary lymphedema. (Liu et al., 2008)
- The low efficiency of naked plasmid technique
Viral vectors for VEGF-C delivery
VEGF-C isoform (VEGF-C156S)
- Saaristo et al. developed a VEGFR3 specific VEGF-C isoform (VEGF-C156S)
- Since VEGF-C activation of VEGFR2 results in blood vessel proliferation and tissue edema.
(Saaristo et al., 2002)
VEGF-C156S using an adenoviral vector to a mouse model of primary lymphedema
- Improved lymphangiogenesis without blood vascular effects
- Development of adeno-associated viral vectors (AAV) for VEGF-C delivery since AAV vectors
- Elicit decreased host inflammatory responses
- Mediate longer-term delivery of the construct compared with adenoviral vectors. (Lai et al., 2002)
Administration of AAV-VEGF-C and VEGF-C vectors to the site of surgical injury
- Increases regeneration and differentiation of lymphatic vessels
- After lymph node dissection in porcine and mouse models.
(Tammela et al., 2007; Lähteenvuo et al., 2011)
- www.frontiersin.org/articles/10.3389/fphar.2022.828513/full
Adenoviral VEGF-C either in the lymph node or in the tissues surrounding it
- Increases regrowth of lymphatic vessels
- Preserves the architecture of the lymph node
- Compared with controls. (Honkonen et al., 2013)
- VEGF-C delivery
- May be helpful for patients treated with autologous VLNT—a surgical treatment for lymphedema.
- Lymphatic regeneration following VLNT
- Is associated with increased VEGF-C expression
- Injections of recombinant VEGF-C increase lymphatic regeneration
- Reconnection in autologous transplanted lymph node fragments. (Sommer et al., 2012; Schindewolffs et al., 2014; Huang et al., 2016)
- www.frontiersin.org/articles/10.3389/fphar.2022.828513/full
Lymfactin®
Phase I and phase II clinical trials
- Adenoviral type 5-based gene therapy vector
- Expresses human VEGF-C. (Hartiala et al., 2020a)
- In 15 patients with breast cancer-related lymphedema who underwent lymph node transplantation and lymfactin injections of various doses (NCT02994771)
- no dose-limiting toxicities 1-year follow-up
- Drug was well tolerated
- Two patients developed infections in the lymphedema limb requiring hospitalization
Phase 2 double-blind, randomized, placebo-controlled, multicenter clinical trial
- 39 patients with 12-months follow-up
- Abstract published in Cancer Research Communications (NCT03658967) (Hartiala et al., 2020b);
- Inconclusive in a company press report. (Herantis Pharma Plc, 2021)
- Drug was generally safe and well-tolerated
- www.frontiersin.org/articles/10.3389/fphar.2022.828513/full
mRNA-based gene delivery techniques
- Nucleoside-modified mRNA encapsulated in lipid nanoparticles (LNPs)
- Safe and effective in various preclinical models. (Pardi et al., 2018; Magadum et al., 2019; Sahu et al., 2019; Polack et al., 2020; Baden et al., 2021; Skowronski and De Serres, 2021; Wang, 2021)
- (Brown et al., 2020)
Nucleoside modified VEGF-C mRNA
- Delivered using lipid nanoparticles
Experimentally induced lymphedema. (Szőke et al., 2021)
- To promote organ-specific lymphangiogenesis and treat
- Injection of the VEGF lipid nanoparticles increased VEGF-C expression locally
- For as long as 20 days after treatment.
- This increased VEGF-C production significantly increased lymphangiogenesis that persisted even 60 days after injection.
- Intradermal injections did not induce significant blood vessel proliferation locally or in other organ systems
- Important since adenoviruses and AAVs can increase gene expression in other organ systems after systemic absorption.
- Single VEGF-C mRNA lipid nanoparticle injection
- Significantly increased edema formation
- Increased adipose deposition in a mouse hindlimb model of lymphedema
VEGF-C expression
- Is increased in lymphedematous tissues (and serum)
- May contribute to increased interstitial fluid accumulation. (Jensen et al., 2015)
- Increased concentration of VEGF-C might bye counterbalanced by
Anti-lymphangiogenic cytokines can directly decrease responsiveness of lymphatic endothelial cells to VEGF-C thus decreasing the efficacy of exogenous VEGF-C delivery for the treatment of lymphedema. (Savetsky et al., 2015; Shin et al., 2015; Ogata et al., 2016) This hypothesis is supported by the finding that VEGF-C overexpression induces transient lymphatic hyperplasia but does not improve lymphatic function in the mouse tail model of lymphedema. (Goldman et al., 2005) It is also possible that over-expression of VEGF-C increases inflammation and blood vessel leakiness thus contributing to increased swelling. (Gousopoulos et al., 2017) Taken together, these findings suggest that the pathophysiology of lymphedema is more complex than a simple deficiency of VEGF-C (or other lymphangiogenic cytokines) and may explain why surgical interventions that decrease scarring or improve lymphatic function are necessary as adjuncts to exogenous VEGF-C delivery for optimal results.
Another potential concern regarding VEGF-C therapy for patients with lymphedema or those at risk of developing the disease, is that VEGF-C is a key regulator of tumor cell growth and metastasis in a variety of solid tumors, including breast cancer. (Skobe et al., 2001; Chen et al., 2012; Kong et al., 2021) Increased VEGF-C expression increases regional lymph node and distant metastasis and is associated with decreased overall prognosis. (Hoshida et al., 2006; Lohela et al., 2009; Tammela and Alitalo, 2010) VEGF-C also promotes proliferation, migration, and invasion of epithelial breast cancer cells. (Karkkainen et al., 2001; Kong et al., 2021) Thus, the delivery of large doses of VEGF-C at the time of surgery to prevent lymphedema development may impact the oncologic aspects of patient care and requires careful study.Other Lymphangiogenic Growth Factors
Other lymphangiogenic growth factors have also been used in preclinical models. For example, fibroblast growth factor 2 (FGF2), promotes lymphangiogenesis by inducing the expression of VEGF-C and VEGF-D. (Javerzat et al., 2002) Onishi et al. demonstrated that topical FGF2 reduces edema, increases lymphatic vessel density, improves lymphatic function (evaluated as the fluorescence intensity of indocyanine green every 3 days), and upregulates VEGF-C expression in a rat tail model of lymphedema. (Onishi et al., 2014)
Hepatocyte growth factor (HGF) regulates migration, proliferation, and differentiation of a wide variety of cells. (Nakamura et al., 1989) HGF is also a powerful promoter of lymphangiogenesis and can act directly on LECs by interacting with its high-affinity receptor-mesenchymal-epithelial transition factor (c-MET). (Kajiya et al., 2005; Cao et al., 2006; Saito et al., 2006) HGF/MET mutations have been identified in patients with primary lymphedema, and single nucleotide polymorphisms of this gene are associated with an increased risk of developing BCRL. (Finegold et al., 2008) MET somatic activating mutations have also been identified in lymphovenous malformations. (Palmieri et al., 2021) Experimental studies have also shown that HGF may have some utility in treating lymphedema. For example, weekly HGF gene therapy decreased swelling and increased lymphangiogenesis in a rat tail model of lymphedema. (Saito et al., 2006)
Retinoic acid agonists such as 9-cis retinoic acid (RA)--a metabolite of vitamin A that is FDA-approved for treating Kaposi’s sarcoma and chronic eczema--increase migration and differentiation of LECs and lymphangiogenesis in vitro and preclinical models of lymphedema. (Choi et al., 2012; Wong, 2021) 9-cis RA treatment also effectively prevented the development of postsurgical lymphedema in a mouse hindlimb model of lymphedema by promoting the formation of collateral lymphatics by activating FGFR signaling. (Jin et al., 2012; Bramos et al., 2016; Perrault et al., 2019; Daneshgaran et al., 2020)
Despite the exciting developments in bench-to-bedside research throughout the past decade, it is important to address the gaps in current clinical trials, including relatively small patient cohorts and lack of standardized and reproducible outcome measures. These challenges and shortcomings are nicely reviewed in other manuscripts. (Forte et al., 2019f; Herantis Pharma Plc, 2021; Walker et al., 2021)Anti-Inflammatory Treatments for LymphedemaInnate Immune Pathways
SOS1
Mutace
- Noonan syndrome 1
- Few with lymphedema (63% with lymphedema)
- Pentrance Medium
- Gain of function
- AD – [55]
SOX18 (SRY-box 18)
Funkce
- Hepatocyte growth factor
- High affinity hepatocyte growth factor receptor (HGF/MET)
- Transcription factor
- Important role in early blood vessel modeling [Downes et al., 2009]
- Differentiation of lymphatic endothelial progenitor cells from venous precursors [Francois et al., 2008]
- VEGFR-3 expression is controlled by PROX1
- A crucial transcription factor for initiation of lymphangiogenesis
- PROX1 is under the control of transcription factor, SOX18 [5]
- Sox18 is expressed in endothelial cells, hair and feather follicles, and the heart
- But no cardiac phenotype is seen in patients or mice [5]
- Cannot activate PROX1 alone
- Needs cooperation from COUP-TFII
- Is expressed throughout the venous system (Srinivasan et al., 2007)
- SOX18 expression appears to be needed only for the initiation of LEC specification [41]
Recessive and dominant mutations in SOX18
- 3 published SOX18 mutations are localized in the
- DNA-binding domain (recessive)
- Transactivation domain (truncating/dominant) [Irrthum et al., 2003] [4]
Mutations SOX18
- Primary lymphedema, congenital lymphedema
- Lymphedema/lymphangiectasia,
- And breast cancer-associated secondary lymphedema
- Hypotrichosis-lymphedema-telangiectasia syndrome (HLTS, OMIM 607823) [4]
- Reduced body hair
- Absence of eyelashes and eyebrows
- Localized cutaneous telangiectasias [4]
- Hypotrichosis-lymphedema-telangiectasia syndrome (HLTS, 607823) [41]
- Hypotrichosis-lymphedema-telangiectasia-renal defect syndrome (HLTRS, 137940) [41]
Sox18-/- mice
- Only minor coat defect [Pennisi et al., 2000a] [4]
- Likely due to redundancy with two close homologs, SOX7 and SOX17
Spontaneous ragged mutant of Sox18 has
- Defective lymphatic and cardiovascular tissues
- Hair follicle defects [Pennisi et al., 2000b] [4]
Dominant nonsense mutation located in the transactivation domain of SOX18
- May compete for DNA binding without transcriptional activation of target genes
- Human mutations may therefore have dominant-negative effects, via competitive transcription factor binding [5]
Two recessive substitutions in the DNA-binding domain
- Likely have less affinity to their promoter binding motifs [5]
Ragged mice Sox18 mutations
- Phenotypically similar [5]
Ovlivnění lymfedému
- HGF/MET pathway is causal for a broad range of lymphedema phenotypes
- new target for the prevention and/or treatment of lymphedema
Expression of HGF via plasmid transfer
- Improves lymphedema via promotion of lymphangiogenesis [2]
Stimulace syntézy SOX18
- ?
Synergické působení
- ?
Inhibice blokátoru
- ?
Agonisté jeho receptoru v lymf. cévách
- ?
SYK-SLP-76 signalling pathway in blood cells (trombocytes)
Syndecan-4
- (Johns et al., 2016)
- Co-receptors for VEGFR-3
- Can enrich the effective cell surface concentration of VEGF-C
- Stabilize the receptor interaction of VEGF-C and render VEGFR-3 signaling pressure-sensitive (Planas-Paz et al., 2012)
TIE receptors
- ANGPT (angiopoetin)-TIE mediated signals or ANG/TIE system
- Important signaling component
- Controlling endothelial cell behavior in angiogenesis and lymphangiogenesis [41]
- Primary open-angle glaucoma (POAG) is often caused by elevated intraocular pressure (IOP)
- Due to increased resistance to aqueous humor outflow (AHO)
- Aqueous humor flows through
- Schlemm’s canal (SC), a lymphatic-like vessel encircling the cornea,
- Intercellular spaces of ciliary muscle cells [43]
- Signaling between angiopoietin (Angpt) and the Angpt receptor Tie2
- Is critical for SC formation
- Indispensable for maintaining SC integrity during adulthood [43]
TIE receptor expression
- Mostly restricted to endothelial cells
- Notable exception of some hematopoietic cells (Batard et al., 1996)
- No ligand has been identified for TIE1
- Considered to be an orphan receptor
- Contributes to signaling in concert with TIE2 [41]
TIE2 receptor
- For all known angiopoietins [41]
Tie2 Expression on Macrophages
- Is Required for Blood Vessel Reconstruction and Tumor Relapse after Chemotherapy [45]
- Hypoxia induced Tie2 expression [45]
- Tie2 prevented macrophages from apoptosis in stress conditions via the AKT-dependent signaling pathway [45]
deletion of Tie2 expression in macrophages
- Significantly prohibited blood supply and regrowth of tumors [45]
Tie2 deleted mice
- no apparent effect on lymphatic vasculature is observed in the (Shen et al., 2014) [41]
- Tie1(?ICD/?ICD) mice
- Appeared normal at E10.5
- Subcutaneous edema by E13.5
- Lymph sac formation occurred in Tie1(?ICD/?ICD) mice
- Defects with the remodeling of primary lymphatic network to form collecting vessels and valvulogenesis
- Abnormal lymphangiogenesis and valve formation in Tie1-ICD(iUCKO/-) mice. [53]
Tie2 deletion in pericytes
- Results in a pronounced pro-angiogenic effect leading to enhanced tumour growth [44]
- Bidirectional, reciprocal EC-pericyte model of Tie2 signalling [44]
Mutace TIE2
- Dominantly inherited venous malformations (600195) [41]
- Primary congenital glaucoma3E (617272) [41]
genetic deletion of Tie2 (Tie2(iUCKO/-)) in neonate mice
- Did not affect lymphatic vessel growth and maturation [53]
Genetic targeting of Tie1
- Embryonically lethal
- Shows lymphatic abnormalities resulting in edema which arises from dysregulated lymph sac formation (D’Amico et al., 2010)
- Defects in
- Lymphatic vessel remodeling
- Collecting vessel formation
- Valve morphogenesis (Shen et al., 2014; Qu et al., 2015) [41]
Tie1 ablation in mice
- Is well tolerated (D’Amico et al., 2014) [41]
Deletion of Angpt1/Angpt2 or Tie2 in adult mice
- Severely impaired SC integrity and transcytosis leading to
- Elevated IOP,
- Retinal neuron damage,
- Impairment of retinal ganglion cell function [41]
- SC integrity is maintained by interconnected and coordinated functions of
- Angpt-Tie2 signaling,
- AHO,
- Prox1 activity
- Functions diminish in the SC during aging
- Leading to impaired integrity and transcytosis [41]
Tie2 reactivation using a Tie2 agonistic antibody
- Rescued the POAG phenotype in Angpt1/Angpt2-deficient mice
- Rejuvenated the SC in aged mice [41]
- Angpt-Tie2 system is essential for SC integrity
- Impairment of this system underlies POAG-associated pathogenesis
- Tie2 agonists could be a therapeutic option for glaucoma [41]
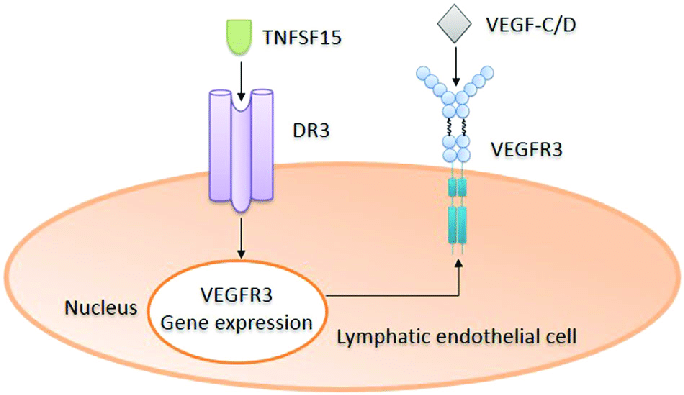
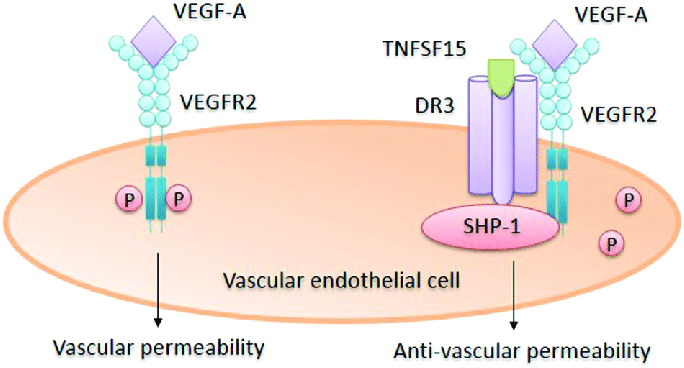
TNFSF15
- Can simultaneously downregulate membrane-bound VEGFR1 and upregulate soluble VEGFR1
- Changing VEGF/VEGFR1 signals from pro-angiogenic to anti-angiogenic [40]
- TNFSF15 inhibits VEGF-induced VEGFR2 phosphorylation
- Curbing VEGFR2-mediated enhancement of vascular permeability
- TNFSF15 is capable of stimulating VEGFR3 gene expression in lymphatic endothelial cells
- Thus augmenting VEGF-C/D-VEGFR3-facilitated lymphangiogenesis [40]
Thrombospondin type 1
Transdiferentiation from venous endothelium
V3 integrin
- VEGFR-2 signalling can be enhanced by interaction (Soldi et al., 1999)
VEGF-A
- Induced Lymphatic hyperplasia (Nagy et al., 2002; Wirzenius et al., 2007) [41]
- VEGFA-induced lymphangiogenesis might also be
- Indirectly mediated by upregulating VEGF-C expression
- In BECs (Skobe et al., 1999; Skobe and Detmar, 2000) or
- In macrophages (Harvey and Gordon, 2012)
- Can be recruited e.g. by VEGF-A via VEGFR-1 (Hiratsuka et al., 1998) [41]
VEGFC-and VEGFD-producing macrophages
(VEGF)-C or D
- Predominantly bind to VEGF receptor-3 (VEGFR-3)
- Promote several downstream signaling pathways regulating lymphangiogenesis
- LEC survival,
- LEC proliferation,
- LEC migration
- Tube formation
- Depends upon the activation of the VEGF-C/VEGFR-3 axis [38]
VEGF-C
- Primary lymphangiogenic growth factor
- Primary ligand that activates VEGFR-3
- Sprouting of endothelial cells from the embryonic veins is crucially dependent on VEGF-C [41]
- A potent inducer of lymphatic endothelial cell proliferation and lymphangiogenesis
- One of the platelet factors secreted from alpha-granules
- VEGFC can also activate the vascular endothelial growth factor receptor 2 (VEGFR2) after proteolysis
- VEGFR2 is only expressed by LEC progenitors
- Only this promotes lymphatic vessel enlargement, not sprouting (Schulte-Merker et al., 2011)
- www.researchgate.net/publication/260485353_Development_of_the_mammalian_lymphatic_vasculature
- VEGFC can act as a morphogen for the progenitor cells of the lymphatic system:
- It induces their differentiation during lymphangiogenesis
- (VEGF)-C-overexpressing human melanomas
- VEGF-C expression also resulted in
- Enhanced tumor angiogenesis,
- Coordinated regulation of lymphangiogenesis and angiogenesis in melanoma progression
- Chemotaxis of macrophages
- VEGF-C and its receptor - VEG FR3
- Key lymphangiogenic growth factor pathway [35]
Exprese VEGF-C
- First produced in larger amounts in regions juxtaposed to the prospective locations of lymphatic sprouting
- Mesenchyme around the developing metanephros
- In the jugular area (Karkkainen et al., 2004; Kukk et al., 1996)
- Forms perhaps a gradient, along which the LECs are migrating (Jha et al., 2017; Yang and Oliver, 2014)
- VEGF-C forms a subgroup within the protein family of vascular endothelial growth factors [41]
VEGF-C activation
- Requires CCBE1 and ADAMTS3
- Disintegrin and metalloproteinase with thrombospondin motifs 3 (ADAMTS3) proteolytic activation of pro-VEGF-C
- Resulting in the mature, active VEGF-C (Jeltsch et al., 2014) [41]
- Before it can signal, it needs to be proteolytically activated by an
Extracellular protein complex comprised of
- Collagen and calcium binding EGF domains 1 (CCBE1) protein
- Protease A disintegrin
- Metallopeptidase with thrombospondin type 1 motif 3 (ADAMTS3) [40]
VEGF-C/VEGFR-3 signaling a ostatní tkáně !
- Lymphatics and VEGF-C expression can be found in almost all tissues during development
- Not all VEGF-C/VEGFR-3 signaling targets endothelial cells
- Brain development, [41]
- Neuronal progenitor cells in the olfactory bulb
- Glial precursor cells in the optic nerve respond to VEGF-C exposure with proliferation (Le Bras et al., 2006) [41]
- VEGF-C signaling in adult mice appears to be able to stimulate neurogenesis (Han et al., 2015) [41]
- In zebrafish, VEGF-C appears crucial for motor neuron axon growth (Kwon et al., 2013) [41]
- In the eye, corneal epithelial cells express VEGFR-3 [41]
- It can act as a decoy receptor removing lymphangiogenic and angiogenic factors thereby maintaining avascularity (Cursiefen et al., 2006) [41]
- Few cells of hematopoietic origin express VEGFR-3 and react to VEGF-C [41]
- Hematopoietic stem cells (Fang et al., 2016; Hamada et al., 2000) [41]
- Megakaryocyte precursors (Thiele et al., 2012) [41]
- Expression of VEGFR-3 by [41]
- Corneal dendritic cells (Hamrah et al., 2003) [41]
- By conjunctival cells of the monocyte/macrophage lineage (Hamrah et al., 2004) [41]
- Immune response in the eye [41]
- Macrophages express VEGFR-3, secrete the VEGFR3 ligand VEGF-C [41]
- Skin, macrophages intriguingly appear to regulate the salt balance of body fluids by secreting VEGF-C [41]
- Proposed to regulate the lymphatic volume [41]
- Gateway function between the hyperosmotic interstitium and normosmolar blood (Machnik et al., 2009) [41]
- VEGFR-3 expressed by tumor associated macrophages (Schoppmann et al., 2002) [41]
- VEGF-C reportedly enhances
- Tumor cell metastasis (Su et al., 2006)
- Leukemic cell growth and proliferation (Dias et al., 2002) by signaling through the VEGFR-3 present on the tumor cells [41]
- VEGFR-3 signaling promotes
- Tumor angiogenesis (Tammela et al., 2008)
- Tumor lymphangiogenesis (Mandriota et al., 2001; Karpanen et al., 2001; Skobe et al., 2001) [41]
- Results in increased metastasis [41]
- VEGF-C– dependent stimulation of lymphatic function
- Promoted drainage and antigen clearance during skin inflammation [42]
VEGF-C absence
- Leads to the failure of lymph sac formation and embryonic death (Hagerling et al., 2013; Karkkainen et al., 2004) [41]
A frameshift mutation
- Identified in 1 family
- Resulting in loss-of-function of the mutant allele [Gordon et al., 2013] [4]
Heterozygous VEGF-C deficiency
- Leads neonatally to severe complications due to insufficient lacteal function
- Resulting chylous ascites (Karkkainen et al., 2004) [41]
Mutations in the human VEGFC gene
- Some forms of hereditary lymphedema (Balboa-Beltran et al., 2014; Gordon et al., 2013) [41]
- Hereditary lymphedema type 1D (615907) [41]
Selective targeting of VEGFR-3-dependent lymphangiogenesis
- Can potentially block cancer progression and metastasis [38]
Blocking VEGF-C/D binding to VEGFR3
- Shown to depress allograft CCL21 production
- Down-regulate DC traffic to the spleen
- Attenuate transplant rejection [42]
Overexpression of VEGF-C
- Can induce lymphangiogenesis and disseminate metastatic tumor cells to lymph nodes
- Use of neutralizing antibodies against VEGF-C and VEGFR-3
- Can prevent tumor lymphangiogenesis and lymphatic metastasis [38]
Lymphangiogenic growth factor VEGF-C supplementation
- Introduced as adjuvant therapy to support lymph node transfer
- Growth factor use remains however associated with a risk for
- (systemic) side effects upon improper dosing
- Unresponsiveness of endogenous lymphatic endothelial cells
- Monotherapy does not mimic the multifactorial physiology of lymphatic growth
- Potentially leading to dysfunctional lymphatic vessels
- Need for repeated administration given
- Short life-span
- Limited bioavailability through rapid diffusion [39]
- Vascular leakage, as seen with high VEGF-C doses
- May even promote lymphedema
Transfer of fibrin-binding VEGF-C
- Did not improve re-epithelialisation
- Increased granulation tissue formation
- Increased matrix remodelling in skin wounds
- Independent of blood vessel growth [39]
Adenoviral expression of VEGF-C in preclinical studies of secondary lymphedema
- Seems to increase lymphangiogenesis around the implanted lymph nodes
- Helping them to connect with the lymphatic system [55]
VEGF-D
- Dissimilar twin of VEGF-C
- First described as c-fos-induced growth factor (FIGF) (Orlandini et al., 1996)
- VEGF-C and VEGF-D are produced as pro-proteins
- Require a multistep proteolytic cleavage before they become active [41]
Aktivace
- First (C-terminal) cleavage
- Similarly executed for both VEGF-C and VEGF-D by furin or the proprotein convertases PC5 and PC7 (McColl et al., 2007; Siegfried et al., 2003)
- Constitutive
- Second (N-terminal) cleavage
- Tightly regulated
- Depends on different enzymes for VEGF-C and VEGF-D (Bui et al., 2016) [41]
- VEGF-C and VEGF-D appear similarly lymphangiogenic in a variety of models like
- Transgenic mice (Jeltsch et al., 1997; Veikkola et al., 2001),
- Adenoviral transduction of skeletal muscle (Rissanen et al., 2003)
- CAM assay (Jeltsch et al., 2003; Oh et al., 1997) [41]
- VEGF-D is not activated by ADAMTS3/CCBE1 (Bui et al., 2016; Jeltsch et al., 2014)
- But instead by plasmin
- VEGF-D would rather act during inflammation- or wound-healing associated lymphangiogenesis (Bui et al., 2016) [41]
- Mature human VEGF-D can bind to both human VEGFR-2 and human VEGFR-3
- Mature mouse VEGFD has been reported to bind only to mouse VEGFR-3, but not to mouse VEGFR-2
Delece VEGF-D
- In mice, without appreciable consequences for the lymphatic system during embryogenesis (Baldwin et al., 2005) [41]
- Adult Vegfd-deleted mice present with initial dermal lymphatics of reduced size and functionality [41]
- Role of VEGF-D during adult lymphangiogenesis
- Specifically perhaps during wound healing (Paquet-Fifield et al., 2013) [41]
- VEGF-D shows a stronger and distinct angiogenic effect compared to VEGF-C in animal models (Duong et al., 2014; Leppanen et al., 2011; Song et al., 2007; Rissanen et al., 2003)
- Maximally processed form of VEGF-D [41]
- Does not anymore activate VEGFR-3, but only the angiogenic receptor VEGFR-2 (Leppanen et al., 2011) [41]
- Difference between VEGF-C and VEGF-D [41]
- Diverging role of the N-terminal -helix for receptor binding (Davydova et al., 2016) [41]
Gene therapy methods to increase VEGF-C expression in lymphedema models
Local delivery of naked plasmid VEGF-C DNA in the rabbit ear or mouse tail models
- Increases VEGFR3 expression
- Decreases swelling and fibroadipose tissue deposition
- Improves lymphatic function as assessed using lymphoscintigraphy. (Yoon et al., 2003)
- Similar results were noted by Liu et al. using a rat hindlimb model of secondary lymphedema. (Liu et al., 2008)
- The low efficiency of naked plasmid technique
Viral vectors for VEGF-C delivery
VEGF-C isoform (VEGF-C156S)
- Saaristo et al. developed a VEGFR3 specific VEGF-C isoform (VEGF-C156S)
- Since VEGF-C activation of VEGFR2 results in blood vessel proliferation and tissue edema.
(Saaristo et al., 2002)
VEGF-C156S using an adenoviral vector to a mouse model of primary lymphedema
- Improved lymphangiogenesis without blood vascular effects
- Development of adeno-associated viral vectors (AAV) for VEGF-C delivery since AAV vectors
- Elicit decreased host inflammatory responses
- Mediate longer-term delivery of the construct compared with adenoviral vectors. (Lai et al., 2002)
Administration of AAV-VEGF-C and VEGF-C vectors to the site of surgical injury
- Increases regeneration and differentiation of lymphatic vessels
- After lymph node dissection in porcine and mouse models.
(Tammela et al., 2007; Lähteenvuo et al., 2011)
- www.frontiersin.org/articles/10.3389/fphar.2022.828513/full
Adenoviral VEGF-C either in the lymph node or in the tissues surrounding it
- Increases regrowth of lymphatic vessels
- Preserves the architecture of the lymph node
- Compared with controls. (Honkonen et al., 2013)
- VEGF-C delivery
- May be helpful for patients treated with autologous VLNT—a surgical treatment for lymphedema.
- Lymphatic regeneration following VLNT
- Is associated with increased VEGF-C expression
- Injections of recombinant VEGF-C increase lymphatic regeneration
- Reconnection in autologous transplanted lymph node fragments. (Sommer et al., 2012; Schindewolffs et al., 2014; Huang et al., 2016)
- www.frontiersin.org/articles/10.3389/fphar.2022.828513/full
Lymfactin®
Phase I and phase II clinical trials
- Adenoviral type 5-based gene therapy vector
- Expresses human VEGF-C. (Hartiala et al., 2020a)
- In 15 patients with breast cancer-related lymphedema who underwent lymph node transplantation and lymfactin injections of various doses (NCT02994771)
- no dose-limiting toxicities 1-year follow-up
- Drug was well tolerated
- Two patients developed infections in the lymphedema limb requiring hospitalization
Phase 2 double-blind, randomized, placebo-controlled, multicenter clinical trial
- 39 patients with 12-months follow-up
- Abstract published in Cancer Research Communications (NCT03658967) (Hartiala et al., 2020b);
- Inconclusive in a company press report. (Herantis Pharma Plc, 2021)
- Drug was generally safe and well-tolerated
- www.frontiersin.org/articles/10.3389/fphar.2022.828513/full
mRNA-based gene delivery techniques
- Nucleoside-modified mRNA encapsulated in lipid nanoparticles (LNPs)
- Safe and effective in various preclinical models. (Pardi et al., 2018; Magadum et al., 2019; Sahu et al., 2019; Polack et al., 2020; Baden et al., 2021; Skowronski and De Serres, 2021; Wang, 2021)
- (Brown et al., 2020)
Nucleoside modified VEGF-C mRNA
- Delivered using lipid nanoparticles
Experimentally induced lymphedema. (Szőke et al., 2021)
- To promote organ-specific lymphangiogenesis and treat
- Injection of the VEGF lipid nanoparticles increased VEGF-C expression locally
- For as long as 20 days after treatment.
- This increased VEGF-C production significantly increased lymphangiogenesis that persisted even 60 days after injection.
- Intradermal injections did not induce significant blood vessel proliferation locally or in other organ systems
- Important since adenoviruses and AAVs can increase gene expression in other organ systems after systemic absorption.
- Single VEGF-C mRNA lipid nanoparticle injection
- Significantly increased edema formation
- Increased adipose deposition in a mouse hindlimb model of lymphedema
- www.frontiersin.org/articles/10.3389/fphar.2022.828513/full
VEGF-C expression
- Is increased in lymphedematous tissues (and serum)
- May contribute to increased interstitial fluid accumulation. (Jensen et al., 2015)
- Increased concentration of VEGF-C might bye counterbalanced by
VEGF-C overexpression
- Induces transient lymphatic hyperplasia
- But does not improve lymphatic function in the mouse tail model of lymphedema. (Goldman et al., 2005)
- Increases inflammation and blood vessel leakiness
- Contributing to increased swelling. (Gousopoulos et al., 2017)
- Pathophysiology of lymphedema is more complex than a simple deficiency of VEGF-C (or other lymphangiogenic cytokines)
- May explain why surgical interventions that decrease scarring or improve lymphatic function
- Are necessary as adjuncts to exogenous VEGF-C delivery for optimal results.
- www.frontiersin.org/articles/10.3389/fphar.2022.828513/full
CAVE: VEGF-C is a key regulator of tumor cell growth and metastasis
- In a variety of solid tumors,
- Including breast cancer. (Skobe et al., 2001; Chen et al., 2012; Kong et al., 2021)
- Increased VEGF-C expression
- Increases regional lymph node and distant metastasis
- Is associated with decreased overall prognosis.
- (Hoshida et al., 2006; Lohela et al., 2009; Tammela and Alitalo, 2010)
- VEGF-C also promotes proliferation, migration, and invasion
- Of epithelial breast cancer cells.
- (Karkkainen et al., 2001; Kong et al., 2021)
- Delivery of large doses of VEGF-C at the time of surgery
- May impact the oncologic aspects of patient care
- www.frontiersin.org/articles/10.3389/fphar.2022.828513/full
VEGFR-2
- Tyrosine kinase receptor
- Mediates most – if not all – functions of the classic hemangiogenic growth factor VEGF-A (Simons et al., 2016)
- Can also be activated by the mature forms of VEGF-C and VEGF-D
- The different forms of VEGF-C and VEGF-D
- Expressed at moderate levels on most LECs
- VEGFA replaces VEGF-C as medium supplement in many LEC culture protocols (Lonza, 2017; PromoCell, 2017) [41]
VEGFR-2-monospecific VEGF-E
- Induced Lymphatic hyperplasia (Wirzenius et al., 2007)
- no increase in lymphatic numbers in VEGF-E overexpressing mice and in mouse ears transduced with a VEGF-A-expressing adenovirus
- VEGFR-2 signalling causes only circumferential growth of lymphatic vessels
- VEGFR-3 signalling causes the generation of new vessels by sprouting lymphangiogenesis [41]
Mutace
- Capillary infantile hemangioma (602089) [41]
VEGFR2
VEGFR2 deletion
- Has little effect on developmental lymphangiogenesis
www.researchgate.net/publication/320024243_Lymphangiogenesis_guidance_by_paracrine_and_pericellular_factors
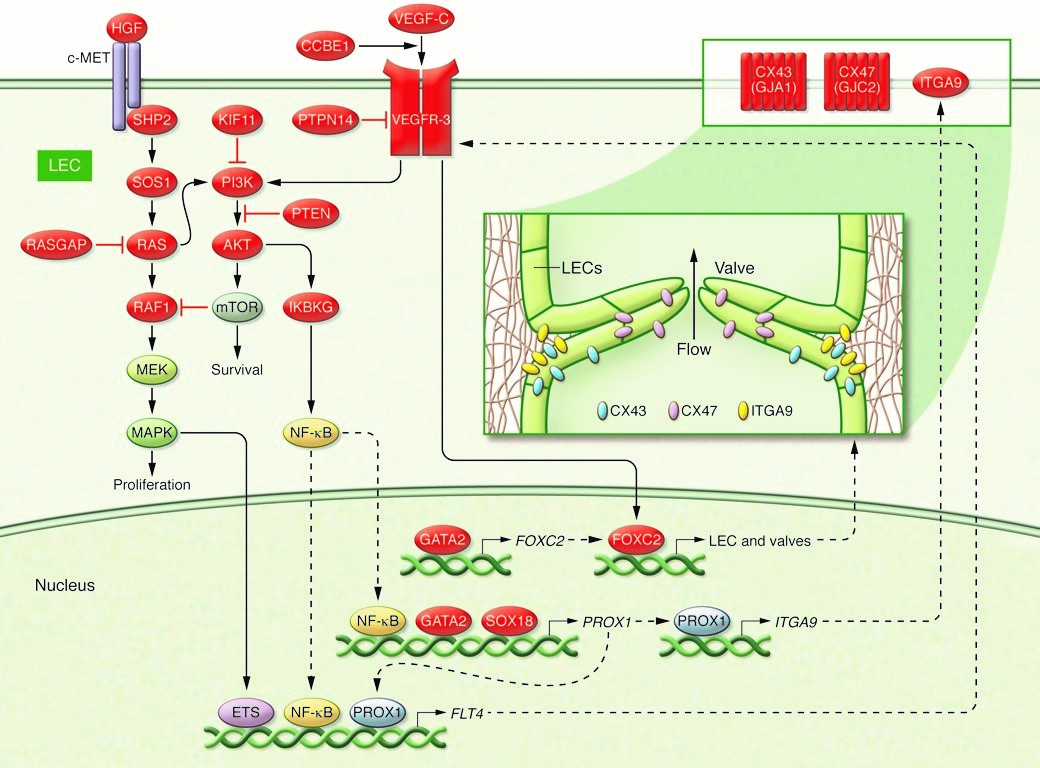
FLT4 gen - VEGFR3 protein
- Transmembrane receptor VEGFR3, encoded by FLT4 pro VEGF-C
- VEGF-C/VEGFR-3 signaling regulate expression of genes essential for lymphatic (and venous) valve development and maintenance. These include
- Gap junction proteins
- Connexins [5]
- Most of the proteins that are encoded by the genes mutated in primary lymphedema
- Seem to act in a single functional pathway involving VEGFR3 signaling
- Important role this pathway in lymphatic development and function [4]
- Regulation of the VEGFR3 signaling pathway
- Central to the molecular mechanism of primary lymphedema [4]
- Key lymphangiogenic growth factor pathway
- Quintessential lymphatic receptor tyrosine kinase [41]
Isoformy in humans
- This diversity has not been seen so far in any other non-primate species (Hughes, 2001) [41]
VEGFR-3s (short isoform)
VEGFR3l (long isoform)
VEGFR-3 expression
- Controlled by PROX1
- A crucial transcription factor for initiation of lymphangiogenesis
- PROX1 is under the control of transcription factor, SOX18 [5]
- SOX18 mutated in the rare hypotrichosis-lymphedema-telangiectasia syndrome (OMIM 607823) [5]
- Variable-onset lymphedema associated with sparse hair and cutaneous telangiectasias [5]
- Declines on BECs during the period of lymphatic budding from venous endothelium and the establishment of the first lymphatic structures
- Later mostly on lymphatics (Kaipainen et al., 1995) with the exception of a few vascular specializations, where it persists into adulthood
- Fenestrated vessels (Partanen et al., 2000)
- Some high endothelial venules (Kaipainen et al., 1995)
- Both LECs and BECs express VEGFR-2 [41]
VEGFR3 signaling

- Theme common to proteins encoded by the 9 genes known to be mutated in patients with primary lymphedema at different levels
- Ligand/receptor (VEGFR3, CCBE1)
- Modification of receptor activity (PTPN14)
- Production of the receptor (SOX18 and GATA2 via PROX1)
- (in)direct control of regulation of transcription (FOXC2, also via GATA2)
- Downstream target gene (GJC2, and FLT4 itself)
- Effects explain more than a quarter of the incidence of primary lymphedema via the
- RAS/MAPK
- PI3K/AKT pathways [5]
- Heterodimerization with VEGFR-2 in response to VEGF-A (Dixelius et al., 2003; Nilsson et al., 2010)
- Could enable VEGFR-3 signaling
- Alternative activation mechanisms
- Involving integrins and mechanoinduction could also play a role (Galvagni et al., 2010; Planas-Paz et al., 2012; Wang et al., 2001) [41]
Mutace FLT4 genu
- Usually an autosomal dominant disorder
- De novo mutations are not infrequent
- Family history of lymphedema is not required for diagnosis
- Family with a particular recessive VEGFR3 mutation has been reported
- Mutations have been discovered in more than 100 families [5]
- VEGFC/VEGFR3 pathway cca 5% lymfedémů [1]
- Type IA hereditary lymphedema (Milroy disease) (153100)
- Capillary infantile hemangioma (602089)
- Most common form of primary lymphedema in humans
- Most of these mutations inactivate the tyrosine kinase activity of VEGFR-3 (Karkkainen et al., 2000)
- In mice, functionally analogous mutations result as well in a lymphedema phenotype (Karkkainen et al., 2001) [41]
Loss of VEGFR-3
- In mice leads to embryonic death
- Due to cardiovascular defects (Dumont et al., 1998) [41]
VEGFR-3 deletion
Deletion of VEGFC and VEGF-D
- In mouse embryos does not, for the most part, affect blood vessels
- With the exception of VEGF-C-induced BEC migration
- Important for the development of the coronary vasculature (Chen et al., 2014a,b) [41]
Cells expressing mutant VEGFR3
- Inhibited autophosphorylation of the receptor
- Indicating downregulation of VEGFR3 signaling [Irrthum et al., 2000; Karkkainen and Petrova, 2000; Ghalamkarpour et al., 2009b] [4]
Klinické projevy mutací
- FLT4 is mutated in Nonne-Milroy disease
- Congenital bilateral lower limb lymphedema
- All reported mutations localized within the 2 intracellular tyrosine kinase domains of the encoded VEGFR3 receptor [4]
- Sometimes present with
- Prenatal pleural effusion
- In utero hydrops
Ovlivnění lymfedemu
- Zvažovali terapii via direct or transgenic delivery of VEGFC [1]
c-Src inhibition
- Would be expected to stabilize VE-cadherin [18]
CAR
- CAR is expressed in platelets
- Platelets play a critical role in the separation between the blood and lymphatic vasculature
- CAR-mediated interactions between lymphatic endothelial cells and platelets
CAR deficiency
- During this critical time window
- Leads to ectopic blood flow in the lymphatic system, edema and hemorrhage
- Due to leaky lymphatic vessels
www.researchgate.net/publication/47755648_Platelets_Covert_Regulators_of_Lymphatic_Development
More differentiated B cell precursors (B220+flk2+)
- Distant from the endosteum
- Closely associated with CXCL12-expressing stromal cells
- Do not express the Notch-1 ligands Jagged-1 or Delta-like-1 (Tokoyoda et al., 2004) [12]
Fibroblast growth factor 2 (FGF2)
- Promotes lymphangiogenesis
- By inducing the expression of VEGF-C and VEGF-D
- (Javerzat et al., 2002)
Topical FGF2
Rat tail model of lymphedema (Onishi et al., 2014)
- Reduces edema,
- Increases lymphatic vessel density,
- Improves lymphatic function
- Evaluated as the fluorescence intensity of indocyanine green every 3 days
- Upregulates VEGF-C expression
- www.frontiersin.org/articles/10.3389/fphar.2022.828513/full
Hepatocyte growth factor (HGF)
- Regulates migration, proliferation, and differentiation (Nakamura et al., 1989)
- HGF is also a powerful promoter of lymphangiogenesis
- Can act directly on LECs
- By interacting with its high-affinity receptor-mesenchymal-epithelial transition factor (c-MET).
- (Kajiya et al., 2005; Cao et al., 2006; Saito et al., 2006)
HGF/MET mutations identified in patients with primary lymphedema
Single nucleotide polymorphisms
- Associated with an increased risk of developing BCRL. (Finegold et al., 2008)
MET somatic activating mutations
- Identified in lymphovenous malformations. (Palmieri et al., 2021)
Rat tail model of lymphedema
- Weekly HGF gene therapy
- Decreased swelling
- Increased lymphangiogenesis (Saito et al., 2006)
- www.frontiersin.org/articles/10.3389/fphar.2022.828513/full
Indukce tvroby venolymfatických anastomóz a chlopní
- Napojení lymfatických cév na okolní žíly
- Low, reversing shear stress is sufficient to induce expression of genes required for lymphatic valve development
- Identified GATA2 as an upstream transcriptional regulator of FOXC2 and the lymphatic valve genetic program [47]
Oscillatory shear stress (OSS)
- Induces the expression of genes, that are important for lymphatic valve development including
- GATA2, FOXC2, GJA4, and ITGA9, in LECs [47]
Notch-1
- Can promote both stem cell self-renewal
- Differentiation toward the lymphoid lineage (Calvi et al., 2003, Pui et al., 1999, Radtke et al., 1999) [12]
p57/CDKN1C genes
- LECs showed enhanced cell proliferation [48]
Retinoic acid agonists
9-cis retinoic acid (RA)
- A metabolite of vitamin A
- FDA-approved for treating Kaposi’s sarcoma and chronic eczema
In vitro and preclinical models of lymphedema
- Increase migration and differentiation of LECs and lymphangiogenesis
- (Choi et al., 2012; Wong, 2021)
Mouse hindlimb model of lymphedema
- 9-cis RA treatment effectively prevented the development of postsurgical lymphedemaby
- Promoting the formation of collateral lymphatics
- By activating FGFR signaling
- (Jin et al., 2012; Bramos et al., 2016; Perrault et al., 2019; Daneshgaran et al., 2020)
- www.frontiersin.org/articles/10.3389/fphar.2022.828513/full
Sympathetic innervation of LNs
- Is well documented
Platelet aggregation is induced
- Is thought to induce 'sealing' and physical separation of the two vascular compartments
- Process depends on the interaction between Podoplanin in LECs and Clec-2 in platelets
Platelet microparticles (PMPs)
- Small membrane-coated vesicles that are released from the plasma membrane upon platelet activation
- Enhance the angiogenic potential of early outgrowth (endothelial progenitor) cells
- Also to stimulate tumor angiogenesis
- Platelets act on lymphatic endothelial cells
- Key regulators of lymphangiogenesis
Po poranění si ruští fighteři nechávali již před 20 a více lety injikovat tzv. "autohemo" - tedy vlastní krev do místa poranění svalů a šlach. I když látky vypuštěné z trombocytů aktivují zároveň i bolest, právě různé růstové faktory z nich stimulují pevnější tvorbu vaziva a urychlují následně srůstání tkání a hojení. Tedy pokud nedojde k zakrvácení přímo do kloubu, neí potřeba mít z nějaké běžné modřiny velkou hrůzu - hraje tam svoji roli v regeneraci - vyjímkou by opět mohly být příliš velké podlitiny - kapsy krve, které naopak mohou představovat riziko vzniku infekce nebo lokálního zvazivovatění. Hned mne napadá otázka - pokud tedy mám pacienta se sekundárním lymfedémem po nějaké operaci - nikoliv pro zhoubné nádorové onemocnění a potřebujeme nastimulovat novotvorbu lymfatických cév - co by se stalo, kdyby mu bylo do podkoží pod místem, kde byly přerušeny lymfatické cévy - injikovány injekce vlastní krve - tedy uměle vytvořené modřiny. Vedlo by to k rychlejší novotvorbě lymfatických cév ? Nezhoršil by se lymfedém ? vlastní krve má na podobné věci každý dost a skoro nic by to nestálo.
V těchto případěch bychom možná mohli lokálně poodat i čistý růstový VEGF-C ?? Existuje již v podobě pro klinické použití? Novotvorba lymfaticých cév by konečně vedla k definitivnímu uzdravení pacientů. Tím by se výrzaně zlepšila kvlait ajejich života a žádná další terapie by již nebyla potřeba.
Tumor-associated lymphopoesis
- Vessel cooption
- Sprouting from the preexisting lymphatics of adjacent tissues
Turbulent flow at vessel branch
- Points within a primitive vascular plexus
- Induces a FOXC2 and GATA2- dependent LEC quiescence and valve endothelial phenotype [47]
Monosomy X - Turner syndrome/nuchal translucency
- Přes 100 případů
- Penetrance Medium [55]
Sex-linked (X0 mice)