nemoci-sympt/BAKTERIALNI-INFEKCE/chlamydie/reakce-na-infekci
AIFM1 AIFM1 – apoptosis-inducing factor, mitochondrion-associated, 1
- C. pneumoniae-induced apoptosis in human coronary artery endothelial cells
- Caspase-independent
- Regulated by Bax and apoptosis-inducing factor (Schöier et al, 2006) [153]
ABCA1 ATP-binding cassette
- Sub-family A (ABC1), member 1 :
- C. pneumoniae modifies cholesterol homoeostasis in human THP-1 macrophages via
- JNK-PPARG dependent pathways
- Downregulation of ABCA1 and ABCG1 (Liu et al, 2010) [153]
ABCG1 ATP-binding cassette
- Sub-family G (WHITE), member 1 :
- C. pneumoniae modifies cholesterol homoeostasis in human THP-1 macrophages via
- JNK-PPARG dependent pathways
- Downregulation of ABCA1 and ABCG1 (Liu et al, 2010) [153]
ACADM acyl-CoA dehydrogenase
- C-4 to C-12 straight chain: ACADM and CPT1A mRNA
- Hypertriglyceridemia produced by C.Pneumoniae was associated with
- Reduced expression of
- Hepatic carnitine palmitoyltransferase-1a (Cpt1a)
- Medium chain acyl-Coenzyme A dehydrogenase (Acadm) (Marangoni et al, 2015) [153]
ACAT1 acetyl-CoA acetyltransferase 1:
- The bacterium induces macrophage related foam cell formation via ACAT1 upregulation (He et al, 2009) [153]
AKT1 Phosphorylation
- Increased by infection in Jurkat cells (Hirai et al, 2012 ) [153]
ALOX12 – arachidonate 12-lipoxygenase
- Platelet activation by C.Pneumoniae is antagonised by 12-lipoxygenase inhibition
- Involved in reactive oxygen species production and oxidation of LDL (Kälvegren et al, 2007) [153]
APOB
- C.Pneumoniae elementary bodies binds to APOB containing fractions of lipoproteins (Bashmakov et al, 2010) [153]
APP amyloid beta (A4) precursor protein
- C.Pneumoniae infection increases brain beta-amyloid deposition in mice (Little et al, 2004) [153]
ATF2 activating transcription factor 2
- During Chlamydia pneumoniae infection, the expression and activity of AP-1 family proteins c-Jun, c-Fos, and ATF-2 were regulated in a time- and dose-dependent manner (Krämer et al, 2015) [153]
ATP sytéza
- Infection stimulates ATP synthes
- Peak of ATP levels occurring mid their infection cycle - max. proliferation [34]
- Sufficient ATP synthesized in the infected cells
- ATP concentration increases above normal levels
- 2–3 x více pro glykogen a glutamát
- Peaked uprostřed inf. cyklu [34]
- + synt. glutamátu via Citr. cyklus [34]
- + synt. laktátu [34]
- + ukládání glykogenu [34]
ATP6V1H ATPase, H+ transporting, lysosomal 50/57kDa, V1 subunit H
- Inhibition inhibits growth (Ouellette et al, 2011 ) [153]
BAD BCL2-associated agonist of cell death
- Disappear during cellular infection (Fischer et al, 2004) [153]
BBC3 BCL2 binding component 3 (Puma) Bim, Puma and Bad proteins
- Disappear during cellular infection (Fischer et al, 2004) [153]
BCL2L11 BCL2-like 11 (apoptosis facilitator) (Bim)
- Bim, Puma and Bad proteins disappear during cellular infection [153]
BIRC2 baculoviral IAP repeat-containing 2
- Regulates immunity to pulmonary infection (Prakash et al, 2009) [153]
BIRC3 baculoviral IAP repeat containing 3:
- Induces the expression induced by infection in a human monocytic cell line (Wahl et al, 2003) [153]
BSG
- EMMPRIN; basigin (OK bloodk group)
- Activated by C.Pneumoniae in monocytes (Schmidt et al, 2006) [153]
Bad proteins
- Disappear during cellular infection (Fischer et al, 2004) [153]
Bim
- Disappear during cellular infection (Fischer et al, 2004) [153]
CALR - Calreticulin
- University of AArhus 2D database
CANX Calnexin
- CPn0809 and CPn1020 colocalise with calnexin (Muller et al, 2008) [153]
CAPRIN2
- Caprin family member 2
- Interacts with the C.pneumoniae inclusion membrane protein Cpn1027 (Flores and Zhong, 2015) [153]
CASP1
- Involved in cleavage of IL1B induced by C.Pneumoniae infection (He et al, 2010) [153]
CASP3
- Chlamydia inhibits apoptosis by blocking caspase 3 activation and mitochondrial cytochrome C release (Airenne et al, 2002) [153]
CASP8
- Activated by infection in T lymphocytes (Olivares-Zavaleta et al, 2011) [153]
CASP9
- Activated by infection in T lymphocytes (Olivares-Zavaleta et al, 2011) [153]
CAT
- Catalase Activity increased by infection in macrophages (Azenabor et al, 2003) [153]
CAV1
- Caveolins associate with Chlamydial inclusion bodies (Webley et al, 2004 ) [153]
CAV2
- Caveolins associate with Chlamydial inclusion bodies (Webley et al, 2004) [153]
CCL5 -chemokine (C-C motif) ligand 5L (RANTES)
- Induced by C. pneumoniae infection in mouse fibroblasts (Chiliveru et al, 2010) [153]
CCND3 Cyclin D3
- Protein levels downregulated by infection in Jurkat cells (Hirai et al, 2012) [153]
CCT5
- Chaperon in containing TCP1, subunit 5 (epsilon)
- University of AArhus 2D database [153]
CD3
- Expression in lymphocyte Molt-4 cells decreased by C. pneumoniae infection (Yamaguchi et al, 2008) [153]
CD40 and CD86 expression
- Downregulated by bacterial MOMP protein in mouse macrophages (Bermudez-Fajardo et al, 2011) [153]
CD69 and CD40LG (cd154 in mouse)
- Downregulated by bacterial MOMP protein in mouse activated CD4(+) T cells (Bermudez-Fajardo et al, 2011) [153]
CDH2 – cadherin 2, type 1, N-cadherin (neuronal)
- C. pneumoniae infection of human brain microvascular endothelial cells resulted increases expression of the
- Beta-catenin
- N-cadherin
- VE-cadherin
- Decreases expression of the
- Tight junctional protein occludin (MacIntyre et al, 2002) [153]
CDH5 – cadherin 5, type 2 (vascular endothelium)
- ( MacIntyre et al, 2002) [153]
CDKN1A – cyclin-dependent kinase inhibitor 1A (p21, Cip1)
- Promoter activated by infection in epithelial cells (Gencay et al, 2003) [153]
CEBPB CCAAT/enhancer binding protein (C/EBP), beta
- C. pneumoniae activates C/EBP-beta, NF-kappaB, and the glucocorticoid receptor in infected epithelial cells (Gencay et al, 2003) [153]
CFL1 Cofilin
- CPn0572 stabilizes pre-assembled F-actin
- By displacing and/or inhibiting binding of the actin-severing protein cofilin (Zrieg et al, 2017) [153]
CHUK – conserved helix-loop-helix ubiquitous kinase
- Activated by C.Pneumoniae infection in in human monocytic lineage cells (Donath et al, 2002) [153]
CKB Creatine kinase B-type
- University of AArhus 2D database [153]
COL1A1 collagen, type I, alpha 1
- Expression decreased by infection in fibroblasts (Baumert et al, 2009) [153]
- Infection of human fibroblasts and smooth muscle cells by C. pneumoniae downregulated type I and III collagen and fibronectin expression (Baumert et al, 2009) [153]
COL3A1 collagen, type III, alpha 1
- Expression decreased by infection in fibroblasts (Baumert et al, 2009) [153]
COL4A3BP collagen, type IV, alpha 3 (Goodpasture antigen) binding protein
- Recruited to the chlamydial inclusion (Moorhead et al, 2010) [153]
COX1 cytochrome c oxidase subunit I
- Macrophages stimulated with C. pneumoniae exhibit early release of reactive oxygen species
- Via up-regulation of NADPH oxidase and cytochrome c oxidase (Azenabor et al, 2005) [153]
CPT1A carnitine palmitoyltransferase 1A - ACADM and CPT1A mRNA
- Hypertriglyceridemia produced by C.Pneumoniae was associated with reduced expression of
- Hepatic carnitine palmitoyltransferase-1a (Cpt1a)
- Medium chain acyl-Coenzyme A dehydrogenase (Acadm) (Marangoni et al, 2015) [153]
CREB3 cAMP responsive element binding protein 3
CRP, pentraxin-related
- Acute phase protein upregulated by this and other infections (de Maat and Kluft, 2001 ) [153]
CSF2 colony stimulating factor 2 (granulocyte-macrophage)
- Intra-nasal bacterial inoculation increases plasma levels of
- IL2, IL5, IL6, IL10, IL12, GM-CSF (= CSF2) , IFN-gamma, and serum amyloid (A Campbell et al, 2010) [153]
CTGF – connective tissue growth factor
- Infection in HeLa cells inceased CTGF, IL6, IL8, IL11, LIF, EGR1 and ETV4 mRNA expression (Peters et al, 2005) [153]
Expression CTNNB1 – catenin (cadherin-associated protein), beta 1, 88kDa
- C. pneumoniae infection of human brain microvascular endothelial cells resulted in increased expression of the
- Beta-catenin,
- N-cadherin,
- VE-cadherin
- Decreases expression of the
- Tight junctional protein occludin (MacIntyre et al, 2002)
CXCL10 – chemokine (C-X-C motif) ligand 10
- Induced by C. pneumoniae infection in mouse fibroblasts (Chiliveru et al, 2010) [153]
CYBA – cytochrome b-245, alpha polypeptide
- Expression increased by infection in porcine coronary arteries
- Increased production of superoxide (Deniset et al, 2012) [153]
CYBB cytochrome b-245, beta polypeptide
- MRNA expression increased by infection of vascular smooth muscle cells (Rivera et al, 2011) [153]
CYCS cytochrome c, somatic:
- Chlamydia inhibits apoptosis by blocking caspase 3 activation and mitochondrial cytochrome C release (Airenne et al, 2002) [153]
CYP7A1
- C. pneumoniae and HCMV infection decrease CYP7A1 promoter activity in human hepatoblastoma HepG2 cells (Michelini et al, 2012) [153]
Calcium channels - L-type voltage sensitive calcium channels
- Influence C.Pneumoniae infection effects in macrophages (Azenabor et al, 2004) [153]
- L-type calcium channel subunits on macrophages include CACNA1C , CACNB1 and CACNB2 (Das et al, 2009) [153]
The copper (Cu)/zinc (Zn) ratio in serum
- Marker of infection/inflammation
- Increased on day 4 and tended to increase again on day 40 in C.p. infected mice [175]
DEFB4A defensin, beta 4A
- Induced by C.Pneumoniae in brain capillary endothelial cells (Tiszlavicz et al, 2011.) [153]
EDN1 endothelin 1
- Expresssion induced by infection in vascular smooth muscle cells (Kern et al, 2009) [153]
EDNRA endothelin receptor type A
- Upregulated by infection of endothelial cells (Kern et al, 2009) [153]
EEA1 – early endosome antigen 1
- C.Pneumoniae colocalizes with
- Surfactant protein (SP)-A-mediated endocytosed lipid
- Early endosomes (EEA1- and Rab5-positive) after 3 and 6 h of infection in pneumocytes (Wissel et al, 2003) [153]
EEF1B2 eukaryotic translation elongation factor 1 beta 2
- University of AArhus 2D database [153]
EGR1 early growth response 1
- Activated by infection of human coronary artery smooth muscle cells (Rupp et al, 2005) [153]
EIF3I eukaryotic translation initiation factor 3, subunit I
- University of AArhus 2D database [153]
EIF5A eukaryotic translation initiation factor 5A
- University of AArhus 2D database [153]
ELK1 – ELK1, member of ETS oncogene family
- C pneumoniae-stimulates phosphorylation of ERK1/2 and Elk-1 in mouse macrophages (Bea et al, 2003) [153]
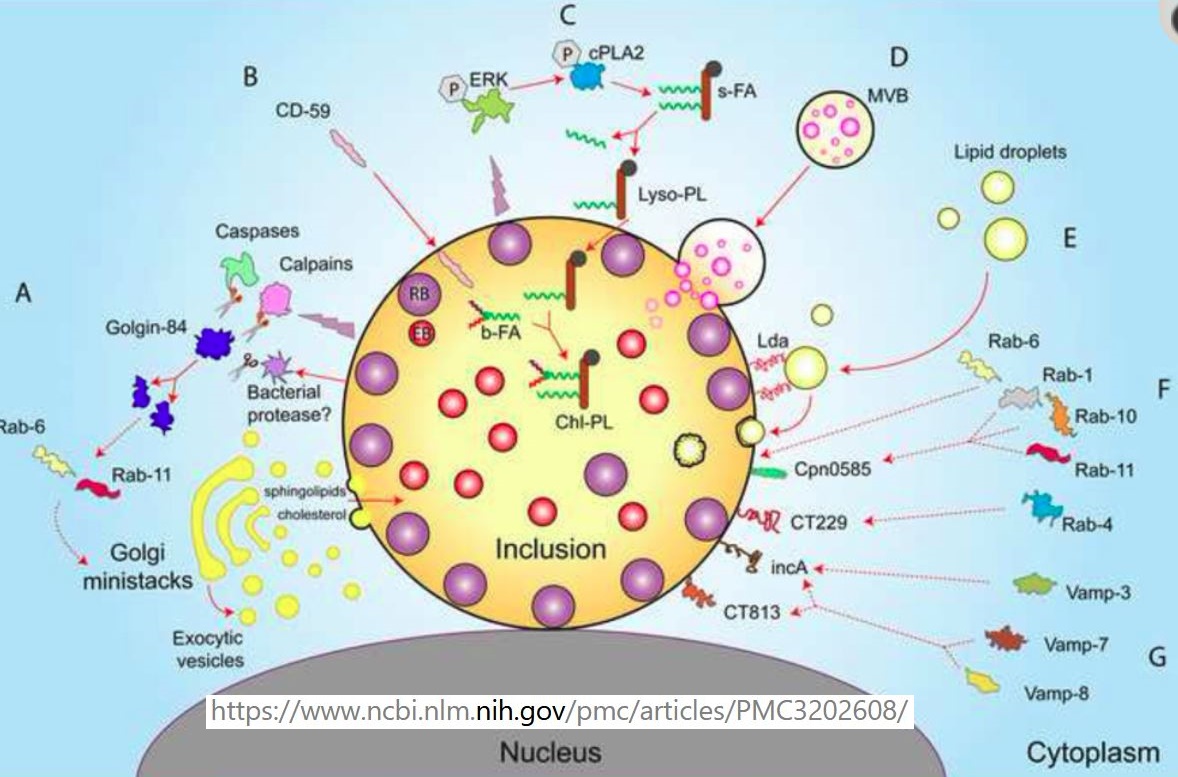
ERK aktivace
- Infection leads to increased phosphorylation of extracellular signal-regulated map kinase (ERK)
- Activates calcium-dependent cytosolic phospholipase A2 (cPLA2)
- CPLA2 removes straight chain fatty acids from the sn2 position of host glycerophospholipids
- To generate lyso-phospholipids (Lyso-PL)
- Chlamydia-derived branched fatty acids (b-FA) are incorporated into Lyso-PL
- To generate Chlamydia-modified phospholipids (Chl-PL) [173]
ETV4 – ets variant 4
- Infection in HeLa cells inceased CTGF, IL6, IL8, IL11, LIF, EGR1 and ETV4 mRNA expression (Peters et al, 2005) [153]
Epitelial cell sensing of pathogenes
- Epithelial cells change their activation status
- Transiently release professional immune cells from inhibition (Mayer & Dalpke, 2007)
- Inhibition is achieved by soluble factors
- Transforming growth factor-beta
- Adaptive regulatory T cells
- Activate professional antigen-presenting cells
- Epithelium-derived factors
- IL-1
- IL-6
- Both are stimulated upon C. pneumoniae infection (Kaukoranta-Tolvanen et al., 1996, Rasmussen et al., 1997)
- Epithelial cells play a role in chlamydial pathogenesis [132]
Epithelial cells
- Aberrant chlamydial bodies can recover
- Generate infectious chlamydial EBs upon removal of the growth-inhibitory factors [13]
F3 F3 – coagulation factor III (thromboplastin, tissue factor)
- Tissue factor, PAI-1, and IL6 expression increased in infected Human vascular endothelial and smooth muscle cells
- NF-kappaB was activated and IkappaBalpha degraded (Dechend et al, 1999) [153]
FAS Fas (TNF receptor superfamily, member 6)
- Chlamydia trachomatis and C.Pneumoniae inhibit fasf-mediated apoptosis (Fischer et al, 2004) [153]
FCGR2A – Fc fragment of IgG, low affinity IIa, receptor (CD32)
- Induced by C.pneumoniae infection of human aortic endothelial cells (Vielma et al, 2002) [153]
FCGR2B – Fc fragment of IgG, low affinity IIb, receptor (CD32)
- [153]
FGA – fibrinogen alpha chain (complexes with FGB and FGG)
- Production incresaed by infection in hepatocytes (Bouwman et al, 2004) [153]
FGF2 fibroblast growth factor 2 (basic)
- Expression upregulated by infection in human endothelial cells (Prochnau et al, 2004) [153]
FLOT1 flotillin 1
- Colocalizes with the Inclusion membrane protein A (IncA) in C. pneumoniae inclusion membranes (Korhonen et al, 2012) [153]
FN1 fibronectin 1
- Expression decreased by infection in fibroblasts (Baumert et al, 2009) [153]
GBP2 – guanylate binding protein 2, interferon-inducible
- Upregulated by infection in mediastinal lymph node lymphocytes (Kyläniemi et al, 2009) [153]
GCLC – glutamate-cysteine ligase, catalytic subunit
- Inhibition increases chlamydial cHSP-60 gene and protein expression in macrophages (Azenabor et al, 2006) [153]
GNLY – granulysin
- Upregulated in T cells by bacterial infection (Walch et al, 2009) [153]
GSK3B glycogen synthase kinase 3 beta
- Interacts with the C.pneumoniae inclusion membrane protein Cpn1027 (Flores and Zhong, 2015) [153]
GSTP1 glutathione S-transferase pi 1
- University of AArhus 2D database [153]
GZMB – granzyme B (granzyme 2, cytotoxic T-lymphocyte-associated serine esterase 1)
- Upregulated in T cells by bacterial infection (Walch et al, 2009) [153]
HAMP – hepcidin antimicrobial peptide
- Peak of viable bacteria in the infected liver associates with increased
- SLC11A2
- Iron contents
- Hepcidin expression (Edvinsson et al, 2008) [153]
HDC – histidine decarboxylase
- Induced by lung infection (Burián et al,2003) [153]
HIF1A hypoxia inducible factor 1, alpha subunit
- Degraded by the the chlamydial protease-like activity factor CPAF
- Which is secreted into the cytoplasm of infected cells (Rupp et al, 2007) [153]
HLA-A major histocompatibility complex, class I, A
- Some infected patients have CD8(+) T cells capable of recognizing the Chlamydial HSP-70 (HLA-A*0201) (Carralot et al, 2005) [153]
HMGB1 – high mobility group box 1
- Released from human aortic endothelial cells suffering aponecrosis following infection (Marino et al, 2008) [153]
HNRNPD – heterogeneous nuclear ribonucleoprotein D (AU-rich element RNA binding protein 1, 37kDa)
- Chlamydial GroEL1 increases VCAM-1 mRNA stability and cytosolic AU-binding factor 1 (AUF1/HNRNPD) levels in endothelial cells (Huang et al, 2013.) [153]
HSPA5 heat shock 70kDa protein 5 (Bip or GRP78 - glucose-regulated protein, 78kDa)
- University of AArhus 2D database [153]
HSPA8 heat shock 70kDa protein 8
- University of AArhus 2D database [153]
HSPA9 heat shock 70kDa protein 9 (mortalin)
- University of AArhus 2D database [153]
HSPD1 heat shock 60kDa protein 1 (chaperonin)
- University of AArhus 2D database [153]
IFIT1 interferon-induced protein with tetratricopeptide repeats 1
- Induced by C. pneumoniae infection in mouse fibroblasts (Chiliveru et al, 2010) [153]
IFIT3 – interferon-induced protein with tetratricopeptide repeats 3
- Upregulated by infection in mediastinal lymph node lymphocytes (Kyläniemi et al, 2009) [153]
IFN
- Ch. pneumonia actively suppresses type I IFN induction [7]
- Preventing phosphorylation and nuclear translocation of IRF3 [7]
- Levels of inhibition corresponded closely to the number of infected cells [7]
- Key signaling molecule in multiple transduction pathways
- Deficiency in this adapter molecule impairs
- IFN-alfa/IFN-ß induction by TLR3, TLR4, TLR7/8, TLR9, RIG-I, and MDA5 [7]
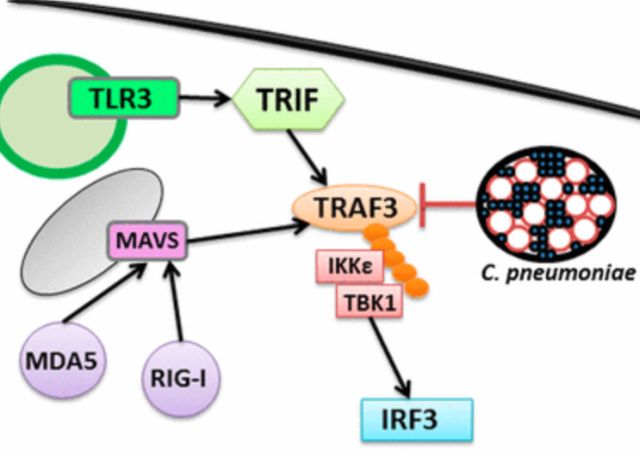
- C. pneumoniae inhibits induction of IFN-ß upon MDA5 and TLR3 engagement [7]
IFNA1
- Infection of murine bone marrow-derived macrophages with Chlamydia pneumoniae induces
- IFN-alpha/beta-
- NF-kappaBeta-dependent expression of IFN-gamma (Trumstedt et al, 2007) [153]
IFNB1 interferon, beta 1, fibroblast
- Infection of human endothelial cells with C. pneumoniae-induced production of IFN-beta (Buss et al, 2010) [153]
IKBKB – inhibitor of kappa light polypeptide gene enhancer in B-cells, kinase beta
- Reduced expression in isolated C.pneumoniae positive human atherosclerotic plaques (Ja et al, 2011) [153]
IL10 interleukin 10
- Regulates intramacrophage Chlamydia pneumoniae replication
- By modulating intracellular iron homeostasis (Bellmann-Weiler et al, 2012.) [153]
IL11 Interleukin 11
- Infection in HeLa cells inceased CTGF, IL6, IL8, IL11, LIF, EGR1 and ETV4 mRNA expression (Peters et al, 2005) [153]
IL12A interleukin 12A (natural killer cell stimulatory factor 1, cytotoxic lymphocyte maturation factor 1, p35) (heterodimer of IL12A and IL12B)
- Intra-nasal bacterial inoculation increases plasma levels of IL2, IL5, IL6, IL10, IL12, GM-CSF, IFN-gamma, and serum amyloid A (Campbell et al, 2010) [153]
IL12B interleukin 12B (natural killer cell stimulatory factor 2, cytotoxic lymphocyte maturation factor 2, p40)
- Intra-nasal bacterial inoculation increases plasma levels of IL2, IL5, IL6, IL10, IL12, GM-CSF, IFN-gamma, and serum amyloid A (Campbell et al, 2010) [153]
IL13 interleukin 13
- Increases the cytokine response to C.Pneumonia in human bronchial epithelial cells (Park et al, 2010) [153]
IL17A interleukin 17A
- C. pneumoniae infection in mice induced IL-17A and IL-17F from day 2 after infection
- Reinfection leads to an incriease in IL25 (aka IL17E) expression (Mosolygó et al, 2013) [153]
IL18 interleukin 18
- C. pneumoniae stimulated IL18 production via MyD88-dependent pathway (Netea et al, 2004) [153]
IL2 interleukin 2
- Intra-nasal bacterial inoculation increases plasma levels of IL2, IL5, IL6, IL10, IL12, GM-CSF, IFN-gamma, and serum amyloid A (Campbell et al, 2010) [153]
IL25 interleukin 25
- C. pneumoniae infection in mice induced IL-17A and IL-17F from day 2 after infection
- Reinfection leads to an incriease in IL25 (aka IL17E) expression (Mosolygó et al, 2013) [153]
IL8 interleukin 8
- Infection in HeLa cells inceased CTGF, IL6, IL8, IL11, LIF, EGR1 and ETV4 mRNA expression (Peters et al, 2005) [153]
IL1B interleukin 1, beta
- Regulates inflammation and fibrosis induced by C.Pneumoniae (He et al, 2010) [153]
IL5 Interleukin 5
- Intra-nasal bacterial inoculation increases plasma levels of IL2, IL5, IL6, IL10, IL12, GM-CSF, IFN-gamma, and serum amyloid A (Campbell et al, 2010) [153]
INSR Insulin receptor
- Downregulated in infected progenitor adipose cells (Shi et al, 2008) [153]
Takže asi přispívá ke vzniku dieabetu II.
IQGAP1 – IQ motif containing GTPase activating protein 1
- C.pneumoniae infection promotes vascular smooth muscle cell adhesion and migration via IQGAP1 (Zhang et al, 2012) [153]
IRAK4 IRAK4, but not IRF3
- Controls C. pneumoniae-induced IFN-alpha and IFN-gamma secretion and bacterial growth in bone marrow macrophages (Trumstedt et al, 2007) [153]
IRF1 – interferon regulatory factor 1
- C. pneumoniae induced the production of low levels of
- Interferon (IFN)-beta
- IRF1
- IRF9 in bronchial and vascular smooth muscle cells (Rödel et al, 2001) [153]
IRF3 interferon regulatory factor 3
- C. pneumoniae infection increases IRF3 and IRF7 nuclear translocation in human vascular endothelial cells (Buss et al, 2010) [153]
IRF7 interferon regulatory factor 7
- C. pneumoniae infection increases IRF3 and IRF7 nuclear translocation in human vascular endothelial cells (Buss et al, 2010) [153]
IRF9 interferon regulatory factor 9
- C. pneumoniae induced the production of low levels of interferon (IFN)-beta and of IRF1 and IRF9 in bronchial and vascular smooth muscle cells (Rödel et al, 2001) [153]
IRS1 insulin receptor substrate 1
- Downregulated in infected progenitor adipose cells (Shi et al, 2008) [153]
ISG15 – ISG15 ubiquitin-like modifier
- Upregulated by IFN- gamma -induced persistent C. pneumoniae infection in HL cells (Mannonen et al, 2007.) [153]
ITGA5 – integrin, alpha 5 (fibronectin receptor, alpha polypeptide)
- Integrin alpha(v)beta(3) involved in C. pneumoniae infection in Hela cells (Caronzolo et al, 2006) [153]
ITGB3 – integrin, beta 3 (platelet glycoprotein IIIa, antigen CD61)
- Integrin alpha(v)beta(3) involved in C. pneumoniae infection in Hela cells (Caronzolo et al, 2006) [153]
ITPR1 – inositol 1,4,5-trisphosphate receptor, type 1
- Expression decreased by infection in porcine coronary arteries
- Also a reducion in L-type calcium channel expression (Deniset et al, 2012) [153]
Protilátky proti chlamidiálnímu HSP 60 - Heat schock protein 60
Human HSP 60
- Anti cHSP 60 zkříženě reagují i proti lidskému HSP60 - vlemi podobný
- Poškozují dál endotel cév - progrese aterosklerozy
Antibodies against Hsp60 (GroEL) (60 kDa) of C. trachomatis
- Important for autoimmune mechanisms in conditions like pelvic inflammatory disease and tubal infertility [124]
Anti HSP 60 bakterií zubního plaku u paradentózy
- Zhoršení progrese aterosklerozy při paradentóze
Infection-induced apoptosis resistance
- Epithelial cells infected with C. pneumoniae
- Resistant to apoptosis induced by treatment with drugs or by death receptor ligation
- Only cells containing large inclusions were protected [161]
- Probably involves mitochondria
- Major integrators of apoptotic signaling
- Mitochondria did not respond to apoptotic stimuli by the release of apoptogenic factors required for the activation of caspases [161]
- Active caspase-3 was absent in infected cells
- Direct modulation of apoptotic pathways in epithelial cells by C. pneumoniae [161]
- Chlamydia-infected host cells are profoundly resistant to apoptosis induced by a wide spectrum of proapoptotic stimuli including
- Kinase inhibitor staurosporine
- The DNA-damaging agent etoposide
- Tumor necrosis factor-alpha
- Fas antibody
- Granzyme B/perforin [162]
- Antiapoptotic activity was dependent on chlamydial but not host protein synthesis [162]
- Inhibited in chlamydia-infected cells
- Activation of the downstream caspase 3
- Cleavage of poly (ADP-ribose) polymerase [162]
- Mitochondrial cytochrome c release into the cytosol induced by proapoptotic factors
- A central step proposed to convert the upstream private pathways into an effector apoptotic pathway for amplification of downstream caspases [162]
Intracellular signaling molecules
- Adaptor proteins
- E3 ubiquitin ligases
- By aktivation of TLR3 or TLR4
- TRIF-dependent K63-linked ubiquitination of TRAF3 [7]
- Crucial for downstream activation of kinases TBK1 and IKK
- Catalyze the phosphorylation of IFN regulatory factor 3 (IRF3)
- Induction of IFN-beta [7]
JUN – jun proto-oncogene
- Infection activates nuclear factor kappaB and activator protein 1 (Jun) in human vascular smooth muscle (Miller et al, 2000) [153]
KNG1 – kininogen 1 (Bradykinin)
- C. pneumoniae infection reduced bradykinin-induced relaxation of precontracted tissue (porcine coronary arteries) (Deniset et al, 2012) [153]
KRT17
- University of AArhus 2D database [153]
KRT18 Keratin
- Degraded by chlamydial CPAF (Savijoki et al, 2008) [153]
KRT8 Keratin
- Degraded by chlamydial CPAF (Savijoki et al, 2008) [153]
LBP – lipopolysaccharide binding protein
- Serum LBP concentration increased post infection in mice (Tiirola et al, 2002) [153]
LCAT – lecithin-cholesterol acyltransferase
- Progressive decline in serum activity post-infection in mice (Tiirola et al, 2002) [153]
LDHA lactate dehydrogenase A
- Released from human aortic endothelial cells suffering aponecrosis following infection (Marino et al, 2008) [153]
LGALS1 lectin, galactoside-binding, soluble
- University of AArhus 2D database [153]
LIF – leukemia inhibitory factor
- Infection in HeLa cells inceased CTGF, IL6, IL8, IL11, LIF, EGR1 and ETV4 mRNA expression (Peters et al, 2005) [153]
LPL Lipoprotein lipase
- Induced by infection in macrophages (Azenabor et al, 2004) [153]
LY96 lymphocyte antigen 96
- Chlamydial cHSP60 induces vascular endothelial and macrophage inflammatory responses
- Via the innate immune receptor complex TLR4/LY96 (Bulut et al, 2002.) [153]
MAP2K1 MAP2K1 – mitogen-activated protein kinase kinase 1 (MEK)
- In epithelial cells C. pneumoniae invasion increased
- MEK-dependent phosphorylation and activation of
- ERK1/2,
- (MAPK1/MAPK3)
- Followed by PI 3-kinase-dependent phosphorylation
- And activation of Akt1 (Coombes and Mahony, 2002) [153]
MAPK8 MAPK8 – mitogen-activated protein kinase 8 (jnk)
- Induction of iNOS by C.pneumoniae requires MyD88-dependent activation of JNK (Rodriguez et al, 2008) [153]
MAVS mitochondrial antiviral signaling protein
- Activated by infection in human endothelial vascular cells (Buss et al, 2010) [153]
MBL2 Mannose binding protein
- Binds to major outer membrane protein on the outer surface of the chlamydial elementary body (Swanson et al, 1998) [153]
MDM2 MDM2 proto-oncogene, E3 ubiquitin protein ligase
- MDM2-mediated degradation of p53 in Chlamydia-infected cells
- Contributes to the anti-apoptotic influence upon the host cell (González et al, 2014.) [153]
MEK/ERK pathway
- Activation essential for chlamydial invasion of HeLa cells [54]
- Sustained activation of Raf-MEK-ERK-cPLA2 required for acquisition of glycerophospholipids and growth by C. pneumoniae [54]
- ERK1/2 kinases
- Essential to initiate the apoptosis delay in C. pneumoniae-infected neutrophils
- Involved in the maintained expression of the anti-apoptotic protein Mcl-1 survival pathways in neutrophils [119]
MMP1 matrix metallopeptidase 1 (interstitial collagenase)
- Expression increased by infection in peripheral blood monocytes (Rupp et al, 2004) [153]
MMP14
- Activated by C.Pneumoniae in monocytes via EMMPRIN (Schmidt et al, 2006) [153]
MMP3
- C. pneumoniae upregulates protein expression of
- Adhesion molecules
- MMP-1
- MMP-3 in vascular cells via OLR1 (Samanta et al, 2017) [153]
MMP7 matrix metallopeptidase 7 (matrilysin, uterine)
- Activated by C.Pneumoniae in monocytes via EMMPRIN (Schmidt et al, 2006) [153]
MMP9 matrix metallopeptidase 9 (gelatinase B, 92kDa gelatinase, 92kDa type IV collagenase)
- Activated by C.Pneumoniae in monocytes via EMMPRIN (Schmidt et al, 2006) [153]
MSR1 macrophage scavenger receptor 1
- Upregulated by infection in human umbilical vein endothelial cells (Shi and Tokunaga, 2004) [153]
MYD88 myeloid differentiation primary response 88
- C.Pneumoniae -induced IL-1beta secretion by macrophages requires TLR2/MyD88 and NLRP3/ASC1/Caspase-1 signaling (Shimada et al, 2011 .) [153]
MYLIP myosin regulatory light chain interacting protein
- Hepatic expression increased in infected mice (Marangoni et al, 2015.) [153]
NF-kappa B–mediated gene activation
- Crucial step in the developmental cycle of C pneumoniae
- Aspirin exerts an anti-chlamydial effect
- Due to the inhibition of C pneumoniae–induced NF-kappa B activation
- Might account for some of the cardioprotective activity of aspirin [20]
- Activated in response to a number of stimuli
- Bacterial lipopolysaccharide
- Phorbol esters
- Interleukin (IL)-1
- Tumor necrosis factor-alfa [20]
- Aspirin/salicylic acid on chlamydial development
- By morphological changes of the inclusions
- Became smaller and less uniformly shaped
Takže indukce persistence ???
- Aspirin at concentrations 10-4 mol/L markedly decreased chlamydial growth
- Serum concentrations of 10-4 to 10-5 mol/L commonly achieved by therapeutic doses of aspirin in humans [20]
- Intracellular growth of C pneumoniae
- Mediated by an activation of the transcription factor NF-kappa B[20]
- Activation of NF-kappa Bconstitutes
- Essential part of the host-pathogen relationship
- Important implications for chlamydial growth [20]
- Treatment with aspirin/salicylic acid
- Resulted in a similar concentration-dependent inhibition of the secretion of IL-6 and IL-8 [20]
- Activation of NF-kappa Bin C pneumoniae–infected macrophages
- Associated with protection of these cells against apoptosis [20]
NFKB1 nuclear factor of kappa light polypeptide gene enhancer in B-cells 1
- C. pneumoniae activates C/EBP-beta, NF-kappaB, and the glucocorticoid receptor in infected epithelial cells (Gencay et al, 2003) [153]
NFKBIA – nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, alpha
- Tissue factor, PAI-1, and IL6 expression increased in infected Human vascular endothelial and smooth muscle cells
- NF-kappaB was activated and IkappaBalpha degraded (Dechend et al, 1999) [153]
- Depleted by C.Pneumoniae infection in in human monocytic lineage cells (Donath et al, 2002.) [153]
NFKBIE – nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, epsilon
- Depleted by C.Pneumoniae infection in in human monocytic lineage cells (Donath et al, 2002.) [153]
NK
- Imunitní reakce směrem k Th1 odpovědi
- Activation of type 1 immune responses essential for protection against chlamydial infection
- Including cell-mediated immunity
- IFN-? production [50]
- NKT-1-like cells
- Predominant IFN-gamma production
- NKT-2-like
- Produce higher IL-4 [50]
Aktivace NLRP3 inflammasome directly
- Mature IL-1ß production
- Critical for proper immune responses against C. pneumoniae infection [7]
NOD-like receptors
- Nod1
- Nod2
- Signal through Rip2 [5]
NOD1 nucleotide-binding oligomerization domain containing 1
- Activated by C.Pneumonia in epithelial cells (Opitz et al, 2005) [153]
NOD2 nucleotide-binding oligomerization domain containing 2
- Activated by C.Pneumonia in epithelial cells (Opitz et al, 2005) [153]
NOS3 nitric oxide synthase 3 (endothelial cell)
- Chlamydia heat shock protein 60 decreases the expression in human and porcine coronary artery endothelial cells (Chen et al, 2009.) [153]
NOX1 NADPH oxidase 1
- Macrophages stimulated with C. pneumoniae exhibit early release of reactive oxygen species via up-regulation of
- NADPH oxidase
- Cytochrome c oxidase (Azenabor et al, 2005) [153]
NR1H3 uclear receptor subfamily 1, group H, member 3 (LXR alpha)
- Involved in bacterial induced foam cell formation (Chen et al, 2008) [153]
NR3C1 nuclear receptor subfamily 3, group C, member 1 (glucocorticoid receptor)
- C. pneumoniae activates
- C/EBP-beta,
- NF-kappaB,
- Glucocorticoid receptor in infected epithelial cells (Gencay et al, 2003) [153]
NRP1 – neuropilin 1
- Upregulated in C. pneumoniae infected human umbilical vein endothelial cells (Shi and Tokunaga, 2004) [153]
Non-Toll-like innate immune proteins
- TLRs work in synergy with the cytosolic NLRs
- Sense bacteria
RIG-I (retinoic acid-inducible gene 1)-like receptors (RLRs)
- Sense viruses
C-type lectin receptors (CLRs)
- Sense fungi (Ishii et al., 2008)
- TLRs and CLRs activate
- Transcription factor NF-kappaB
- MAP kinases
- Expression of many immune and inflammatory genes
- Cytokines and chemokines
NOD1 and NOD2
- Respond to muropeptides derived from bacterial peptidoglycan (McDonald et al., 2005)
- Chlamydial genome encodes enzymes required for peptidoglycan synthesis (Kalman et al., 1999; Stephens et al., 1998)
- Motif needed for NOD recognition
- Present during Chlamydia spp. infection
- C. pneumoniae-infected HEK293 cells
- NOD1- and NOD2-mediated NF-kappaB activation was demonstrated (Opitz et al., 2005)
- NOD1 recognition
- NF-kappaB activation
- Secretion of proinflammatory cytokines during
- In vitro infection with C. trachomatis (and C. muridarum)
- Vaginal infection
- NOD1 deficiency in mice
- Did not influence the course of the infection, cytokine secretion or pathology (Welter-Stahl et al., 2006)
Nalp3, IPAF and NAIP
- Components of the so-called inflammasomes
- Contain caspase-1
- Cleaves pro-IL-1beta and pro-IL-18 into their mature forms (Akira et al., 2006; Ishii et al., 2008)
CLRs
- Mediate endocytosis and/or phagocytosis
- Role in antigen presentation
- Keep endogenous glycoprotein levels constant (Palsson-McDermott & O'Neill, 2007)
- Subset of CLRs, for example, Dectin-1
- Respond to various PAMPs
- Can also act as phagocytic receptors
- Mannose receptor
- Implicated in the entry of C. trachomatis into host cells (Kuo et al., 2002)
- PAMP recognition by CLRs is also potentially connected to the induction of innate response genes (Robinson et al., 2006)
- Triggering receptors expressed on myeloid cells (TREM-1)
- Can increase inflammation
- Can be modified by C. trachomatis infection (Cooper et al., 2008) [132]
OCLN occludin
- C. pneumoniae infection of human brain microvascular endothelial cells resulted increases expression of the
- Beta-catenin,
- N-cadherin,
- VE-cadherin
- Decreases expression of the
- Tight junctional protein occludin (MacIntyre et al, 2002) [153]
OCRL oculocerebrorenal syndrome of Lowe ( phosphatidylinositol-bisphosphatase [EC:3.1.3.36] )
- Recruited to the chlamydial inclusion (Moorhead et al, 2010) [153]
OPTN Optineurin (fip2)
- Rab11-OPTN (Fip2) is recruited to the nascent inclusion upon internalization (Mölleken and Hegemann, 2017) [153]
OSBP oxysterol binding protein
- Recruited to the chlamydial inclusion (Moorhead et al, 2010) [153]
PCNA proliferating cell nuclear antigen
- Expression increased by infection in Human aortic vascular smooth muscle cells
- Associated with decreased nuclear binding of p53 (Selzman et al, 2003.) [153]
PDIA3 protein disulfide isomerase family A, member 3
- University of AArhus 2D database [153]
Prostaglandin E2
- Release increased by infection in lymphocyte Molt-4 cells (Yamaguchi et al, 2008) [153]
PHACTR1 – phosphatase and actin regulator 1 phosphatidylethanolamine
- Chlamydial receptor on HeLa cells, along with asialo-GM1 (aka GA1) and asialo GM2 (Krivan et al, 1991) [153]
PI3K/Akt kinases
- Essential to initiate the apoptosis delay in C. pneumoniae-infected neutrophils
- Involved in the maintained expression of the anti-apoptotic protein Mcl-1
- Leads to the activation of NF-?B-dependent release of IL-8 by infected neutrophils
- Leads to the maintenance of Mcl-1 expression in neutrophils [119]
PI4K2A phosphatidylinositol 4-kinase type 2
- Alpharecruited to the chlamydial inclusion (Moorhead et al, 2010) [153]
PI4K2B phosphatidylinositol 4-kinase type 2 beta
- Recruited to the chlamydial inclusion (Moorhead et al, 2010) [153]
PLA2G7 phospholipase A2, group VII (platelet-activating factor acetylhydrolase, plasma)
- In atherotic plaques isolated from carotid arteries Lp-PLA(2) (PLA2G7) associates with plaque macrophages and C. pneumoniae (Atik et al, 2010) [153]
PLAUR plasminogen activator, urokinase receptor
- Secretion increased by C.Pneumoniae in monocytes via EMMPRIN (Schmidt et al, 2006) [153]
PPARA peroxisome proliferator activated receptor alpha
- C.Pneumoniae induces foam cell formation in macrophages via PPARA and PPARG (Mei et al, 2009) [153]
PPARG peroxisome proliferator-activated receptor gamma
- C.Pneumoniae induces foam cell formation in macrophages via PPARA and PPARG (Mei et al, 2009) [153]
PSMB6 proteasome (prosome, macropain) subunit, beta type, 6
- University of AArhus 2D database [153]
PSMB9 proteasome (prosome, macropain) subunit, beta type, 9 (large multifunctional peptidase 2)
- University of AArhus 2D database [153]
PSMD9 proteasome subunit
- University of AArhus 2D database [153]
PSME1 proteasome (prosome, macropain) activator subunit 1 (PA28 alpha)
- University of AArhus 2D database [153]
PTK2 protein tyrosine kinase 2 ( aka FAK)
- Tyrosine phosphorylation of FAK preceded its appearance in a complex with the p85 subunit of PI 3-kinase ( PIK3R1) during C.pneumoniae invasion (Coombes and Mahony, 2002) [153]
PTK2B protein tyrosine kinase 2 beta (FAK)
- Tyrosine phosphorylation of focal adhesion kinase induced by infection of host cells (Coombes and Mahony,2002) [153]
PYCARD PYD and CARD domain containing (inflammasome component) required for caspase (CASP1) cleavage of IL1B
- Induced by C.Pneumoniae infection (He et al, 2010) [153]
Pathogen-sensing pattern recognition receptors - PRRs
- Present in macrophages and dendritic cells
- Include
- Toll-like receptor (TLR) family
- Present at the cell surface or within phagosomes (Takeda & Akira, 2005)
- Cytosolic nucleotide-binding and olimerization domain (NOD) -like receptor family (NLR) (Carneiro et al., 2007) [135]
- Recognition of PAMPs (pathogen-associated molecular patterns) by PRRs results in
- Intracellular signaling cascades
- Production of inflammatory cytokines
- Upregulation of costimulatory molecules
- Activation of the antimicrobial defense [135]
- PAMPs potentially present in Chlamydia spp. or during the chlamydial developmental cycle include
- Lipopolysaccharide (endotoxin)
- Peptidoglycan (Chopra et al., 1998)
- V.s. lipoprotein
- V.s. bacterial nucleic acid [135]
Phospholipase D - PLD
- TLR4 agonist inducing the expression of
- IL23A, IL6, IL1B, TGFbeta, and CCL20 in monocytes
- CXCL9, CCL20, CCL4, CCL2, ICAM1, and VCAM1 in HUVECs
- Plaque-derived T cells produce IL-17 in response to CpPLD (Benagiano et al, 2012) [153]
Puma
- Disappear during cellular infection (Fischer et al, 2004) [153]
RAB1 RAB1, member RAS oncogene family
- Binds to the Chlamydia pneumoniae inclusion membrane protein Cpn0585 (Cortes et al, 2007) [153]
RAB10
- Binds to the Chlamydia pneumoniae inclusion membrane protein Cpn0585 (Cortes et al, 2007) [153]
RAB11A
- Binds to the Chlamydia pneumoniae inclusion membrane protein Cpn0585 (Cortes et al, 2007) [153]
RAB4A – RAB4A, member RAS oncogene family
- Binds to the Chlamydia pneumoniae inclusion membrane protein Cpn0585 (Cortes et al, 2007) [153]
RAB7A
- After entry, the early chlamydial inclusion acquires early endosomal Rab GTPases including
- Rab4, Rab5, Rab7,
- Rab11, Rab14 (Mölleken and Hegemann, 2017) [153]
RAC1 – ras-related C3 botulinum toxin substrate 1 (rho family, small GTP binding protein Rac1)
- Vascular smooth muscle cell infection increased cell membrane-associated RhoA and Rac1 (Dechend et al, 2003) [153]
RELA v-rel reticuloendotheliosis viral oncogene homolog A (avian)
- Degraded by the chlamydial protease CPAF (Christian et al, 2010) [153]
RFX5 regulatory factor X, 5 (influences HLA class II expression)
- Degraded by the chlamydial protease CPAF (Fan et al, 2002) [153]
RHOA – ras homolog family member A
- Vascular smooth muscle cell infection increased cell membrane-associated RhoA and Rac1 (Dechend et al, 2003) [153]
RPLP2 ribosomal protein, large, P2
- University of AArhus 2D database [153]
RPS6KB1 ribosomal protein S6 kinase, 70kDa, polypeptide 1
- Phosphorylation inhibited by infection in Jurkat cells (Hirai et al, 2012) [153]
SAA1 serum amyloid A1
- Intra-nasal bacterial inoculation increases plasma levels of IL2, IL5, IL6, IL10, IL12, GM-CSF (= CSF2) , IFN-gamma, and serum amyloid (A Campbell et al, 2010) [153]
SELE – selectin E
- Expression increased by infection in human umbilical vein endothelial cells
- Along with ICAM1 and VCAM1 (Kaukoranta-Tolvanen et al, 1996) [153]
SELP – selectin P (granule membrane protein 140kDa, antigen CD62)
- C.pneumoniae binds to platelets and triggers P-selectin expression (Kälvegren et al, 2003) [153]
SERPINE1 – serpin peptidase inhibitor, clade E (nexin, plasminogen activator inhibitor type 1), member 1
- Tissue factor, PAI-1, and IL6 expression increased in infected Human vascular endothelial and smooth muscle cells
- NF-kappaB was activated and IkappaBalpha degraded (Dechend et al, 1999) [153]
SLC11A2 solute carrier family 11 (proton-coupled divalent metal ion transporters), member 2
- Peak of viable bacteria in the infected liver associates with
- Increased SLC11A2
- Iron contents
- Increased hepcidin expression (Edvinsson et al, 2008) [153]
SMAD4 – SMAD family member 4
- Reduced expression in isolated C.pneumoniae positive human atherosclerotic plaques (Ja et al, 2011) [153]
SOD1 superoxide dismutase 1, soluble
- Upregulated by C.Pneumoniae in macrophages (Azenabor and Chaudhry, 2003) [153]
SQSTM1 sequestosome 1
- Down-regulated at 24hrs
- Up-regulated at 72hrs following infection of neuronal cells (Slutter, PhD thesis, 2011) [153]
STAT1
- Regulates IFNB and IFNB control of Chlamydial infection
- Controls IDO1 (Rothfuchs et al, 2006) [153]
STX6 Syntaxin 6
- Recruited to the chlamydial inclusion (Moore et al, 2011) [153]
TGFB1
- C. pneumoniae-infected human bronchial epithelial cell showed enhanced secretion of
- VEGF
- TGF-beta
- TIMP-1 (Park et al, 2010) [153]
Innate and adaptive immunity gene transcripts increased at 48 hours in Chlamydia pneumoniae-infected THP1 cells
- CCL2
- COLEC12
- CYBB
- DEFB4
- DMBT1
- IFNB1
- IL1F5
- IL1F8
- IL1RN
- IL6
- IRAK2
- NFKBIA
- NLRC4
- PTAFR
- SERPINA1
- TLR8
- TNF [153]
TICAM1 – toll-like receptor adaptor molecule 1 (Trif)
- C. pneumoniae facilitates foam cell formation
- Via activation of both MyD88-dependent and MyD88-independent (i.e.TICAM1-dependent and IRF3-dependent) pathways
- Downstream of TLR2 and TLR4 (Chen et al, 2008) [153]
TIMP1 TIMP metallopeptidase inhibitor 1
- C.pneumoniae-infected human bronchial epithelial cell showed enhanced secretion of
- VEGF, TGF-beta, and TIMP-1 (Park et al, 2010) [153]
TLR2 toll-like receptor 2
- Recognises C.Pneumoniae on dendritic cells (Prebeck et al, 2001) [153]
TLR3
- C.pneumoniae initiates signalling via TLR3 and MAP Kinase(s) (Wang et al, 2013) [153]
TLR4 toll-like receptor 4
- Recognises C.Pneumoniae on vascular smooth muscle cells (Sasu et al, 2001) [153]
TLRs
- Best-characterized innate immunity receptors
- 10 functional TLRs identified in humans
- In the cytoplasmic or endosomal membranes
- Sense a variety of microbial ligands at the cell surface
- TLR1, TLR2, TLR4, TLR5, TLR6 and TLR11
- Within endosomes
- TLR3, TLR7, TLR8 and TLR9) (Kawai & Akira, 2005) [132]
Binding of the ligands to TLRs leads to
- Recruitment of adaptor proteins
- Most often myeloid differentiation factor (MyD) 88
- TRIF
- TRAM
- TIRAP [132]
- Adaptor proteins recruit a signaling complex composed of
- IRAKs
- TRAF6 (Kawai & Akira, 2007) [132]
- This leads to activation of
- NF-?B
- protein-1
- Induction of inflammatory cytokine expression, including
- TNF-alpha
- IL-1beta
- IL-6 (Misch & Hawn, 2008) [132]
- Enables interaction with adaptive responses [132]
- Bronchial epithelial cells express functional TLR1–6
- Ligation of a TLR initiates a cascade of signaling pathways
- MyD88-dependent (TLR1, TLR2, TLR4, TLR5, and TLR6)
- TRIF-dependent (TLR3 and TLR4) manner [6]
During Chlamydia spp. infection
- NF-?B is activated
- Several proinflammatory cytokines are produced in various cell types [132]
- Suggesting signaling through PRRs (Kaukoranta-Tolvanen et al., 1996; Rasmussen et al., 1997; Dechend et al., 1999; Kol et al., 1999; Prebeck et al., 2001; Gencay et al., 2003) [132]
- TLRs
- Play a role in the pathogenesis of atherosclerosis (Cook et al., 2004; Misch & Hawn, 2008)
- C. pneumoniae proteins Cpn 0809, Cpn 0980 and OMP2
- Activate mouse macrophages through TLR2 and TLR4 (Jiang et al., 2008)
- Chlamydial lipopeptide exposed on chlamydial EB
- Macrophage infectivity potentiator
- Mediated proinflammatory response in human macrophages through TLR2/TLR1/TLR6 (Bas et al., 2008) [132]
TLR2
- Surface-exposed
- Senses inter alia lipopeptides
- Chlamydiae are gram-negative bacteria and have lipopolysaccharide
- Signaling through TLR2 is often observed
- Live chlamydial elementary bodies (EB)
- Can initiate TLR2-mediated signaling (Rodríguez et al., 2006)
- LPS of C. trachomatis
- Signal via TLR2 (Erridge et al., 2004) [132]
- Induction of early immune response in human lung tissue x C. pneumoniae Droemann et al. (2007)
- Foam cell formation in RAW 264.7 cells Cao et al. (2007)
- Early immune response in mice Rodríguez et al. (2006)
- Chemokine release in vascular smooth muscle cells (Yang et al. 2005)
- Increased ATP content in mouse bone marrow macrophages (Yaraei et al. 2005a)
- Oxidized LDL-induced death augmented in RAW 264.7 cells (Yaraei et al. 2005b)
- Induction of proinflammatory cytokines in PBMC (Netea et al. 2002)
- Activation of mouse bone marrow-derived dendritic cells (Prebeck et al. 2001)
- Production of proinflammatory cytokines in PBMC (Netea et al. 2004)
- CHsp60 - Accumulation of PMN, increased serum levels of chemokines in mice ( Costa et al. 2004)
- CHsp60 Stimulation of mouse bone marrow-derived dendritic cells (Costa et al. 2002)
- Omp2, 0809, 0980 TNF-? production in RAW 264.7 cells (Jiang et al. 2008) [132]
- C. trachomatis
- Colocalizes with the inclusion (O'Connell et al. 2006) [132]
- LPS Activation of NF-?B promoter (Erridge et al. 2004)
- C. muridarum
- Development of oviduct pathology (Darville et al. 2003)
- Secretion of cytokines in oviduct epithelial cells [132]
TLR4
- Senses lipopolysaccharide from gram-negative bacteria (Akira et al., 2006)
- Bacterial lipopolysaccharide
- Classic ligand for TLR4 (Akira et al., 2006)
- Lipid A structures may vary considerably
- All gram-negative bacteria do not synthesize lipopolysaccharide with hexaacyl lipid A
- Chlamydial lipopolysaccharide contains nonhexaacyl lipid A structures
- Might not effectively activate host defenses via TLR4
- Could allow growth of bacteria
- May even promote persistence [132]
- Chlamydial heat shock protein (Hsp)60
- Act as a ligand both for TLR4 and for TLR2 (Bulut et al., 2002; Costa et al., 2002)
- But signal through TLR4 (Maguire et al., 2005) [132]
- Abundant cytoplasmic chaperone molecule
- Also surface exposed on some bacteria (Paju et al., 2000)
- Produced upon interferon (IFN)-gamma treatment during C. pneumoniae infection (Molestina et al., 2002)
- Could thus be accessible to TLRs [132]
- IFN-gamma production in mouse bone marrow macrophages Rothfuchs et al. (2004)
- Expression of tissue factor in RAW 264.7 cells Bea et al. (2003)
- C. pneumonia omp2, 0809, 0980 TNF-alpha production in RAW 264.7 cells Jiang et al. (2008)
- Hsp60
- Activation of human PBMC Maguire et al. (2005)
- Accumulation of PMN, increased serum levels of chemokines in mice da Costa et al. (2004)
- Stimulation of mouse bone marrow-derived dendritic cells Costa et al. (2002)
- C. trachomatis Hsp60
- Induction of apoptosis of human trophoblasts Equils et al. (2006)
- Activation of mouse macrophages and human endothelial cells Bulut et al. (2002) [132]
MyD88
- MyD88-dependent pathways
- Induction of various inflammatory cytokines [6]
- Adapter molecule associated with both TLR2 and TLR4
- Important role in initiating an early effective response against C. pneumoniae and clearance of the bacteria (Rodríguez et al., 2007)
- MyD88-deficient mice
- Severe inflammation with elevated inflammatory cytokine levels
- Due to the defective initial clearance of the bacteria (Naiki et al., 2005) [132]
- Human ectocervical cells, MyD88 was required for IL-8 secretion
- TLR2 and MyD88 were localized to C. trachomatis inclusions
- Suggesting active intracellular signaling through TLR2 (O'Connell et al., 2006) [132]
- C. pneumoniae
- Inflammatory response in mice Rodríguez et al. (2007)
- Bacterial clearance and survival in mice Naiki et al. (2005)
- IFN-gamma production in mouse bone marrow macrophages Rothfuchs et al. (2004)
- IFN-gamma synthesis (through IL-18 release) in PBMC Netea et al. (2004)
- C. trachomatis
- Colocalizes with the inclusion O'Connell et al. (2006)
- Induction of IFN-gamma-inducible protein 10 in mice Nagarajan et al. (2005)
TRIF (TIR domain containing adaptor-inducing IFN-beta)
- Adaptor recruited as a consequence of recognition of double-stranded (ds) RNA by TLR3
- V.s. responsible for IFN-beta production by Chlamydia muridarum-infected oviduct epithelial cells (Derbigny et al., 2007)
- TRIF-dependent pathways
- Induction of type I IFNs [6]
IRAK4
- A part of several TLR-signaling pathways
- Control growth of C. pneumoniae in mouse bone-marrow-derived macrophages
- Through IFN-alpha and IFN-gamma secretion [132]
TRAF6
- Regulated NF-?B activation (Trumstedt et al., 2007) [132]
Inhibice TLRs
Some virulent bacteria
- Can interfere directly with TLR function
- By secreting proteins that inhibit the signaling
- Lead to impaired host defense (Cirl et al., 2008)
- V.s. Stafylokokus aureus, některé bakterie zubního plaku aj. [132]
Chlamydial infection
- Remains to be studied [132]
Variation in TLR genes (SNPs)
- Can confer increased risk of certain infections (Misch & Hawn, 2008)
- Analysis of polymorphisms in genes encoding PPR signaling can elucidate the pathogenesis of C.pneumonia related conditions [132]
TNFSF11 – tumor necrosis factor (ligand) superfamily, member 11 (RANKL)
- IL6 and RANKL expression increased by C.pneumoniae infection in human osteoblast-like cell line hFOB (Bailey et al, 2008) [153]
TP53 tumor protein p53
- PCNA Expression increased by infection in Human aortic vascular smooth muscle cells
- Associated with decreased nuclear binding of p53 (Selzman et al, 2003.) [153]
TRAF3 aktivace
- TNFR-associated factors (TRAFs)
- Signaling molecule
- Crucial for type I IFN production
- Potential target for disarming immune responses
- Consistent with an impairment in inflammatory responses to the microorganism
- Epithelial cells with C. pneumoniae does not lead to IFN-ß production
- Cells are prevented from activating IFN regulatory factor 3
- Mediated by C. pneumoniae–dependent degradation of TRAF3
- Independent of a functional proteasome [7]
- C. pneumoniae expresses a unique protease targeting TRAF3-dependent immune effector mechanisms [7]
TRAF3 TNF receptor-associated factor 3
- Degraded by the bacterium (Wolf and Fields, 2013) [153]
TRAF3IP2 TRAF3 interacting protein 2
- Binds to a secreted chlamydial protein, CP0236 (Wolf et al, 2009) [153]
TUBB Beta tubulin
- Degraded by chlamydial CPAF (Savijoki et al, 2008) [153]
TXN Thioredoxin
- University of AArhus 2D database [153]
Trombocyty
- C pneumoniae, at a chlamydia:platelet ratio of 1:15
- Adheres to platelets and triggers P-selectin expression after 1 minute
- Causes an extensive aggregation and ATP secretion after 20 minutes of incubation
- Inhibition of glycoprotein IIb/IIIa with Arg-Gly-Asp-Ser or abciximab
- Markedly reduced C pneumoniae-induced platelet aggregation
- Exposure of C pneumoniae to polymyxin B abolished the stimulatory effects on platelet activation
- V.s. chlamydial lipopolysaccharide has an active role
- Other tested bacteria had no or only moderate effects on platelet functions
UQCRC1 ubiquinol-cytochrome c reductase core protein I
- University of AArhus 2D database
USF1 upstream transcription factor 1
- Degraded by the chlamydial protease CPAF (Zhong et al, 2001) [153]
USP18 – ubiquitin specific peptidase 18
- Upregulated by infection in mediastinal lymph node lymphocytes (Kyläniemi et al, 2009) [153]
VCAM1
- Chlamydia pneumoniae infection promotes the transmigration of monocytes through human brain endothelial cells
- Via upreculation of
- VCAM1 and ICAM1 on endothelial cells
- Integrins ITGB1, ITGB2, and ITGAM on monocytes (MacIntyre et al, 2003.) [153]
VIM Vimentin
- Degraded by chlamydial CPAF (Savijoki et al, 2008)
ZDHHC17 (HIP14)
- Interacts with bacterial Cpj0783 (Yanatori et al, 2015 ) [153]
Blokáda apoptózy infikovaných buněk
- Jedním z mechanismů, které při infekci C. pneumoniae vedou k chronickému průběhu
- Blokován release of mitochondrial cytochrome c into the cytosol
- Activation of caspase 3
- Essential role in cellular apoptosis [66]
- Hostitelské buňky infikované chlamydiemi jsou zcela rezistentní k apoptóze
- Indukované širokým spektrem proapoptotických stimulů
- Včetně inhibitoru kinázy taurosporinu
- Látky ničící DNA etoposidu
- Tumor necrosis faktoru alfa
- Antiapoptotické působení je pozorováno v pozdním stadiu vývojového cyklu chlamydií
- Závislé na produkci některých jejich bílkovin, které blokují uvolnění
- Mitochondriálního cytochromu c
- A aktivaci kaspázy (proteázy s cysteinem v aktivním místě)
- Antiapoptotický stav buňky infikované C. pneumoniae se zdá bránit T-buněčné destrukci [121]
CD8+ T cells
- Peptide-specific IFN-gamma-secreting CD8+ T cells
- In local and systemic compartments after primary infection
- Expanded after pathogen re-exposure [120]
- Were cytotoxic
- Displayed a memory phenotype
- Secreted IFN-gamma and TNF-alpha, but not IL-4 [120]
- Lysed Cpn-infected macrophages
- Lytic activity was inhibited by brefeldin A
- Cpn peptide-specific CD8+ CTL suppressed chlamydial growth in vitro
- By direct lysis of infected cells
- By secretion of IFN-gamma and other soluble factors [120]
- CD8+ CTL protect against Cpn [120]
CHUK – conserved helix-loop-helix ubiquitous kinase
- Activated by C.Pneumoniae infection in in human monocytic lineage cells (Donath et al, 2002) [153]
Dendritické buňky (DCs) z leukocytů
- Production of tumor necrosis factor (TNF)-alfa in temporal artery biopsies of GCA patients [12]
Immature DC
- Express less major histocompatibility complex (MHC), CD40, CD80, CD83, and CD86 surface molecules [12]
- Uptake and process antigens [12]
Mature DCs
- Poorly phagocytic
- Highly efficient stimulators of naive T-cell responses
- Express the surface molecules mentioned above
- Produce IL-12 and TNF-alfa during the maturation process
- Inflammation
- Differentiation of T-helper type 1 (Th1) cells
- Upregulation of costimulatory molecules [12]
- Failure to fully activate DCs may result in an unbalanced immune response
- Increased infections [12]
- Direct stimulation of DCs by Chlamydiae
- Soluble factors such as TNF-alfa and IL-1 produced by activated DCs and monocytes
- Potent autologous or heterologous inducers of DC maturation [13]
- Chlamydial development in DCs is not exponential as in epithelial cells
- Suppression may be due to activation of an immune response against Chlamydiae [13]
- DCs play a major role in the development of chronic chlamydial infections
- Inclusions within DCs
- Not formed with replicating C. trachomatis and Chlamidophila psittaci
- Appear to be killed within the first few hours postinfection
- Fusion of the inclusion with host cell lysosomes
- Normally inhibited in chlamydia-infected epithelial cells
- C. trachomatis survives and replicates in human DCs
- C. pneumoniae survive in the DCs long after infection
- Chlamydiae are in a persistent form [13]
- May be reactivated in the condition of immunosuppression
- During TNF-alfa inhibition
- Production of TNF-alfa by Chlamydiae-infected DCs
- Contributes to the growth arrest
- Full-bacterial replication could be restored by TNF-alfa inhibition
- Leading to reactivation
- Mature DCs restrict the intracellular growth of many bacteria
- Including Mycobacteria
- Mycobacterium tuberculosis interferes with TLR signaling blocking DC maturation
- Contribute to the chronic carriage of this pathogen.
- C. pneumoniae stimulates DC to secrete cytokines, particularly TNF-alfa
- Leads subsequently to cell maturation
- Inhibition of TNF-alfa
- Reduced cytokine/chemokine production by the DCs
- Beneficial effects of TNF-alfa blockage in inflammatory diseases
- Increased risk of infection or reactivation
- Inhibition of TNF-alfa enhances Chlamydiae progeny in human DC
- Maturation of the DC is crucial for chlamydial growth arrest and persistence [13]
- Inclusions in DCs
- Atypical, polymorphic
- Contain lower numbers
- Aberrant chlamydial bodies
- Indicating persistent/arrested infection [13]
Chlamydia pneumonia infection effect on dendritic cells GDS3753
- Top misregulated:
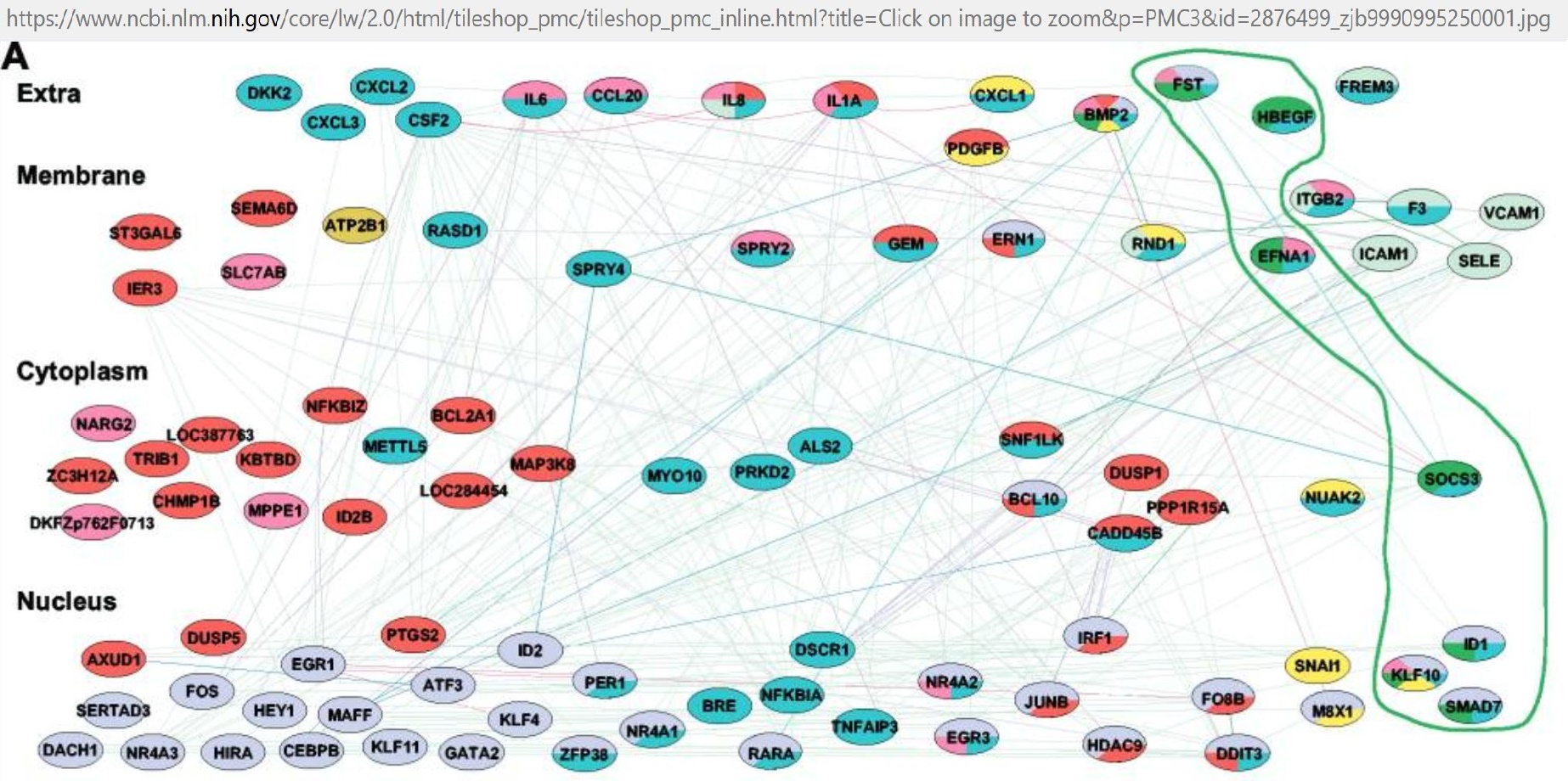
- ACSM5 ACSS1 ACTL9 ACVRL1 ADAD1 ADAMTS6 ADIPOQ AGBL3 AKT2 ALDH5A1 ALOX15B ANP32D APC ARHGEF26 BMS1P20 C15orf48 C3orf79 CA6 CCDC110 CCL20 CCL5 CCL5 CCR7 CD163 CD1E CD69 CD80 CD84 CECR6 CFB CLEC10A CLEC2B CLRN3 CLYBL CNIH4 COL14A1 COX17 CP CPEB1 CSNK1A1P1 CST7 CTLA4 CTNNBIP1 CXCL1 CXCL10 CXCL11 CXCL5 CXCL9 CYB5B DANCR DBR1 DCLK1 DSG2 DUSP4 DUSP5 ECT2 EGR1 EHD1 EIF3A ELAVL1 EPB41L4A ERAP1 ERCC2 EXOSC5 F13A1 FAM47A FAM49A FBXO16 FCAR FOXR2 FRMD4A FRMD4B GBP1 GBP1P1 GBP4 GBP5 GCH1 GGNBP2 GPD1L GPR132 GPR132 GPR157 GREM1 HTR2B IDO1 IFI27 IFIH1 IFITM3 IFT27 IGLC1 IL15RA IL1A IL1B IL1R2 IL23A IL2RA IL2RA IL6 INHBA INSL6 ISG20 ISG20 KANSL1L KLHL31 KLHL7 LILRA5 LINC00705 LINC00877 LINC01191 LINC01289 LINC01521 LITAF LNPEP LOC100129518 SOD2 LOC100130111 LOC100287290 LOC100506082 LOC100653137 LOC101927907 LOC101928927 SNORA9 SNHG15 LOC101929657 LOC101929800 LOC101927424 LOC440896 LOC101930476 LINC00476 LOC102725116 LOC105375839 LOC284513 LOC401220 LOC441666 LPAR6 LRRC25 LRRCC1 LY9 MAGI1 MBNL3 MBOAT1 MDGA2 MEF2C MLLT3 MPP7 MS4A6A MSI2 MT1M NCBP3 NCKAP5 NEMP1 NEURL3 NF2 NRGN NT5DC2 OR2J2 OR2M4 OSMR PDE4B PGAP1 PLGLB2 PLXDC2 PML POSTN PPP2R2A PRAM1 PRAP1 PRELP PRIM2 PRMT2 PSTK RAB7ARAD51L3 RFFL RFPL3S RFTN2 RGS13 RGS18 RIMBP2 RNF144B RORA-AS1 RRAS2 RSAD2 S1PR1 SDS SEPP1 SETD6 SGPP2 SHISA3 SKA3 SLAMF7 SLAMF7 SLAMF7 SLC17A8 SMYD1 SNAI3 SNTG2 SOX17 SPATA4 SRPK2 STAMBP STIM1 STRIP2 STXBP4 STYK1 TAP1 THBS1 TMED5 TMEM37 TNFAIP3 TNFAIP3 TNFAIP6 TNFAIP6 TNFSF4 TRAF1 TRIM4 TRIM69 TTC39A UPB1 VEGFA VMO1 ZDHHC14 ZNF395 ZNF606 ZNF865
- Njau et al, 2009 [153]
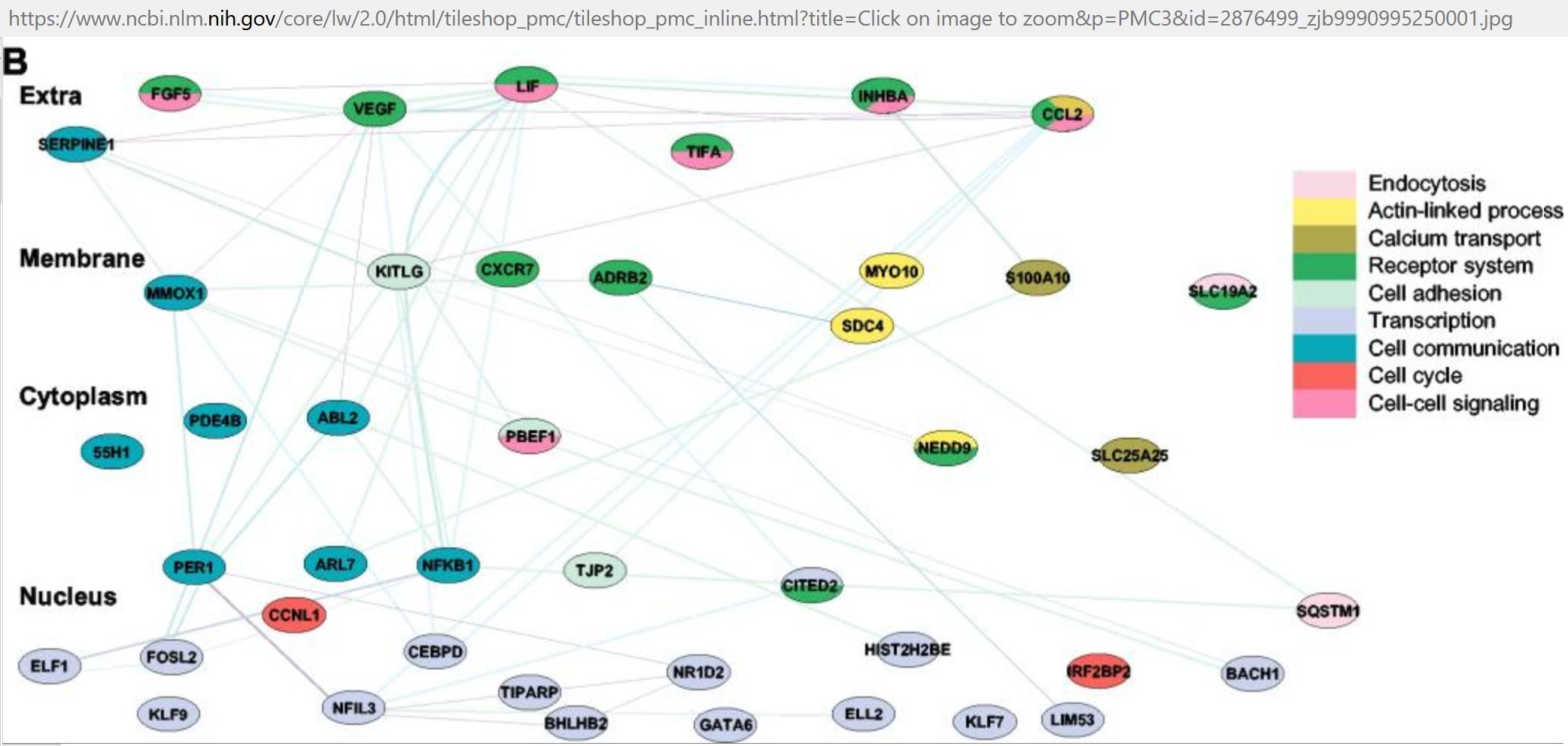
cDNA array analysis of altered gene expression in human endothelial cells in response to Chlamydia pneumoniae infection
- Upregulated
- BDNF CCL2 CXCL12 F2R FGF2 FPR3 GAP43 HBEGF IFNAR1 IGF1R IGFBP4 IL1B IL8 INHBA IRF1 PDAP1 PDGFB PDGFRA TNFAIP6
- (Coombes and Mahony, 2001.) [153]
Infection of U937 Monocytic Cells with Chlamydia pneumoniae Induces Extensive Changes in Host Cell Gene Expression
- Upregulated:
- ADAM9 ANXA1 ANXA5 CCL1 CCL2 CCL3 CCL4 CCL8 CCR1 CCR8 CCRL2 CD44 CD58 CD82 CD83 CD86 CEBPB CREM CSF3 CXCL1 CXCL3 ELK3 EMP1 F5 FABP4 FCER2 FCGR2B GK GPER HBEGF HDAC2 IFNGR2 IL1B IL1RN IL4R IRF8 ITGAX JUNB MMP3 MMP10 MMP12 MMP3 MYB NCOA1 NFATC1 NFKBIA NPC1 NR3C1 PLAU PPBP RGS1 SCMH1 SERPINB7 SMARCD2 SRC TAF5 TANK TFE3 TM4SF1 TMEM2 TNF TNFAIP3 TNFAIP6 TNFSF7 ZNF9
- Downregulated:
- ACAT2 CDC25B CDCA5 CDK2 EEF2 ERP29 FKBP1A HNRNPA1 HPRT1 HSP90AB1 ICSBP1 IMPDH2 KIF23 KIF2C LIG1 LMO2 MCM7 MGP MRPS35 MYB NPM1 NUTF2 PAICS PHB PLK PPAT PRKDC RAB32 RBBP4 RFC1 RNASE2 SCMH1 SNRPD1 SRRM1 TPR
- Virok et al, 2003 [153]
Innate and adaptive immunity gene transcripts
- Increased at 48 hours in Chlamydia pneumoniae (Cpn)-infected THP1 cells
- CCL2 COLEC12 CYBB DEFB4 DMBT1 IFNB1 IL1F5 IL1F8 IL1RN IL6 IRAK2 NFKBIA NLRC4 PTAFR SERPINA1 TLR8 TNF [153]
Increase in the rate of glycolysis [34]
- Not caused by chlamydial metabolic activity but instead is a host cell response to the infection [34]
- + exprese GLUT-1 transport. na vnější mem. [34]
- 2-3x jako po stim. inzulínem [34]
Human flotillin-1 protein
- Ch. pneumonia disrupt flotillin-1-mediated endocytosis
- Bacterial intracellular growth was attenuated in the flotillin-1-silenced cells
- Flotillin-1 colocalized with the inclusion membrane protein A (IncA) in the C. pneumoniae inclusion membranes
- Associated with IncA in detergent-resistant membrane microdomains (DRMs) in biochemical fractioning [8]
Mannose-binding lectin (MBL)
- Complement-activating innate immune defense serum protein
- Binds to mannose and acetylglucosamine sugar groups on different microorganisms
- MBL inhibits infection of HeLa cells by different Chlamydia species
- MBL participates in the protection against C pneumoniae [18]
- MBL variant alleles
- Decreased serum levels of functional MBL
- Associated with an increased risk of respiratory infections especially during early childhood [18]
- Associated with accelerated development of severe atherosclerosis [18]
- MBL inhibits the uptake of the bacteria to target cells in vitro [18]
- Modulate the severity of the infection, rather than protect against [18]
- Might inhibit the dissemination of C pneumoniae from the lungs by monocytes/macrophages to the blood [18]
T lymfocyty
- Zásadní pro eradikaci C.p.
- Patients with scarring sequelae of repeated/persistent C. trachomatis infection
- Depressed cell-mediated immune response
- Indicated by decreased interferon (IFN)-gamma levels
- Increased antibody levels
- Suggesting a predominance of a T-helper (Th)-2-type immune response in these patients [22]
- Cell-mediated immunity may also be deleterious to the host in favouring the development of inflammation [22]
- Even mice that are genetically predisposed to mount a Th2-type response
- In a primary C. pneumoniae infection, elicit a Th1-type response in reinfection
- Markedly increased IFN-gamma production [22]
CD4+ T-cells
- Contributory role
- Especially in the later phases of infection [22]
- In the absence of CD8+ T cells, may even enhance the bacterial growth in the lungs [50]
CD8+ T cells
- CD8+ T-cells crucial role in protection
- Absence of CD8+ T-cells
- Infection appears to progress rapidly [22]
- Prominent role in protective responses
- Clearance of primary infection
- Against reinfection with C. pneumoniae [50]
- CD8+ T cells are the major cell type producing a IFN-gamma
- During the adaptive immune phase of C. pneumoniae infection [50]
- GalCer substantially enhances the level of protective immunity to this infection in mice [50]
- Peptide-specific IFN-gamma-secreting CD8+ T cells
- Local and systemic compartments after primary infection
- Expanded after pathogen re-exposure [59]
- CD8+ T cell lines to the 18 Cpn epitope-bearing peptides (Cpn peptide-specific CD8+ CTL) were:
- Cytotoxic
- Displayed a memory phenotype
- Expressed a CD44high memory phenotype [59]
- Secreted IFN-gamma and TNF-alpha
- But not IL-4 [59]
- Lysed Cpn-infected macrophages
- Inhibited by brefeldin A
- = endogenous processing of CTL Ags [59]
- Suppressed chlamydial growth in vitro
- Direct lysis of infected cells
- Secretion of IFN-gamma and TNF [59]
- Most organisms released by lysed MKF were most likely in the form of noninfectious reticulate bodies
- Human CD8+ T cell-derived granulysin [59]
- Protect against Cpn
- Develop vaccines against acute and chronic Cpn infections [59]
- Ags need to traffic into the cytosol for CD8+ CTL induction [59]
- CD8+ T cells and IFN-? are critical for protection [59]
- Absence of CD8+ T cells [59]
- Cpn-infected mice have increased bacterial burdens and disease severity [59]
- Animals lacking IFN-? signaling
- Bacterial loads are higher
- Clearance of organisms is greatly hampered [59]
- Cpn-derived Ags contributing to MHC class I-restricted CD8+ T cell responses
- Remain unidentified [59]
- Type 1 cytokine secretion
- Effector cells recognize multiple MHC class I-restricted epitopes
- From Cpn Ags endogenously processed
- By productively infected macrophages
- A CD8 zabijí MKF [59]
- Other CD8+ T cell-derived factors (suppress HIV replication):
- RANTES
- Macrophage inflammatory proteins-1alpha and -1beta
- IL-16 [59]
Zkřížená imunizace
- Significant proportion of children display antibodies against C. pneumoniae before the onset of sexual activity and exposure to C. trachomatis STI.
- Comparative analysis of the genomes has revealed a very high degree of synteny between C. pneumoniae and C. trachomatis genomes
- Suggesting the likelihood of several shared antigenic determinants
- Immune responses induced by one pathogen may mediate cross-species protective immunity or pathogenic responses against the other [124]
- Mice vaccinated intranasally with live C. pneumoniae
- But not mock (PBS) immunized animals, displayed
- High levels of splenic cellular antigen-specific IFN-gamma production
- Serum antibody response against C. muridarum and C. trachomatis
- Significant reduction in the vaginal C. muridarum shedding as early as day 12 after secondary i.vag. challenge
- Compared to PBS (mock) immunized mice
- At day 19 after C. muridarum challenge, 100% of C. pneumoniae vaccinated mice had cleared the infection
- Compared to none (0%) of the mock immunized mice
- Which cleared the infection by day 27
- At day 80 after C. muridarum challenge significant reduction in the incidence (50%) and degree of hydrosalpinx
- Compared to mock immunized animals (100%)
- Respiratory C. pneumoniae infection
- Induces accelerated chlamydial clearance and reduction of oviduct pathology following genital C. muridarum challenge
- May have important implications to the C. trachomatis-induced reproductive disease in humans
Potencionální reprodukční výhoda, která mohla pomoct v selekci lidí, co jsou permisivní k chr. C.pneumonia ???
Jak to tak dále čtu, tak v.s. čím silnější imunitní reakce včetně progrese patogenního procesu, tím silnější ochrana proti infekci jiným kmenem... [124]
Infection of the lung
- Characterized by an early phase dominated by
- Granulocytes
- One of the main target of C. pneumoniae infection
- Act as “infecting” cells and reservoir of the pathogen [25]
- Late phase dominated by
- Alveolar macrophages [120]
Respiratory epithelium
- Acts as a mechanical barrier
- Bronchial epithelial cells have tight junctions
- Cells are covered by mucus
- Cells secrete antimicrobial substances
- Cilia clear the airways from microorganisms mechanically
- Epithelium detects invading pathogens
- Pathogen-sensing pattern recognition receptors - PRRs (Bals & Hiemstra, 2004; Hippenstiel et al., 2006; Mayer & Dalpke, 2007) [135]
Infekce myší
Days 5–60 after infection
- Mild interstitial mononuclear cell infiltration
- Perivascular and peribronchiolar lymphoid cuffing
12 days postinfection
- Percentage of CD8+ T cells in lungs from infected mice was almost twice
- Pulmonary bacterial loads commonly peaked 2–4 days before maximal inflammatory responses
- With mean IFUs per lung of 3 × 105 ± 9 × 104 after primary infection [120]
Reinfection - 2 days postinoculation
- Moderate to marked pneumonia
- Lymphoid infiltrate [120]
Reinfection - days 8–12 days postinoculation
- Peaked pneumonia
- Gradually declined
- Remained present for up to 2 months
- Pulmonary bacterial loads commonly peaked 2–4 days before maximal inflammatory responses
- With mean IFUs per lung of 2.9 × 104 ± 8 × 103 [120]
Respirační infekce
- Infikované epiteliální buňky jsou definitivně vyloučeny ze slizničního povrchu a zanikají
- Chlamydie jsou v této pro ně vulnerabilní fázi spojené s přechodem do jiné buňky vnímavé k eliminaci
- U většiny lidí zničeny v průběhu týdnů až měsíce [121]
- Dosaženo nejspíše CD4 Th1-dependentními cytokiny
- Interferon gamma
- Neutralizujícími protilátkami (např. slizniční IgA) [121]
- Důležitou roli v nasměrování adaptivní T-buněčné imunity mají i NKT buňky (natural killer T cells)
- Jejich aktivace infekcí C. pneumoniae indukuje následně fenotypové a funkční změny dendritických buněk významných v rozvoji protektivní imunity
- Za určitých podmínek je C. pneumoniae je schopna pronikat z místa primárního kontaktu hlouběji do organismu [121]
iNOS induction
- Impaired 6 days postinfection
- Pulmonary levels of IFN-? and TNF are elevated
- Induction of iNOS in macrophages upon C. pneumoniae infection
- Controlled by MyD88 via two pathways:
- NF-kappa B activation and phosphorylation of the MAPK JNK
- Nuclear translocation of c-Jun, one of the two components of the AP-1 complex
- Phosphorylation of STAT1 and expression of IFN regulatory factor 1 (IRF-1)
- Delayed in the absence of MyD88 after C. pneumoniae infection [13]
- Optimal induction of iNOS during C. pneumoniae infection is required:
- Action of the MyD88-dependent transcription factors
- NF-kappa B
- AP-1 [13]
- The MyD88-independent transcription factors
- Phosphorylated STAT1
- IRF-1 [13]
Characterization and intracellular localization of putative Chlamydia pneumoniae effector proteins
- () Müller et al, 2008) [153]
- UP IFIT1 IFIT3 TTLL11Down EIF5 GLP2R
Lymphocytes in blood
- Control of this pathogen in these cells may be critical in the development of chronic inflammatory diseases
- Activity of antibiotics against C. pneumoniae in lymphocytes menší než v epiteliích [2]
- C. pneumoniae replicates mainly in macrophage/monocytes and neutrophils
- To a lesser degree, in epithelial cells [5]
- C. pneumoniae in blood cells, such as monocytes as well as lymphocytes, is resistant to antibiotic treatment [53]
- Whereas C. pneumoniae in epithelial cells is not [53]
Macrophage cell line response to Chlamydia pneumoniae GDS2651
- Upregulated:
- ABCB1B ABR ACOD1 ACSL1 ACTG1 ACTR3 AHNAK ALAS1 ANGPTL2 ARHGEF1 ATF3 ATP6VOA1 BCL2A1D BCL3 BCL6 CASP4 CCL2 CCL3 CCL4 CCL7 CDR2 CEBPB CEBPD CITED2 CLEC4E CLIC4 CPD CSTF2 CTNND1 CXCL2 CYTIP DUSP16 EBI3 EHD1 EIF3E ELL2 EPRS ETF1 ETS2 FAM49B FAS FLNB FPR1 GADD45A GCH1 GPR137B GSR GTPBP4 HCLS1 HIGD1A HK2 HMOX1 HNRNPA1 HP HSD17B12 HSD17B7 HSD3B2 HSPA8 IFIH1 IFITM3 IFRD1 IGH2BP2 IL1B IL1RN IMPDH1 KLF6 LILR4B LPL LTB MANF MAPK6 MAPK8 MAPKAPK2 MARCKSL1 MTHFS MTPN MYADM MYO1D NAB1 NEK1 NFKB1 NFKBIA NFKBIZ ODC1 OTUD4 PIM1 PIP1 PIRB PLK2 POU2F2 PSMA3 PSMB6 PSMB7 RFWD2 RGCC RHOC RMDN3 RNF19B RPL7 RSAD2 RTN4 SAA3 SCOC SLC11A2 SLC20A1 SLC25A25 SLC2A1 SLC31A1 SLC4A7 SLFN2 SLPI SOCS3 SOD2 SPATA13 SRSF2 SUSD6 TAF1D TIAL1 TIMM17A TMEM131 TMEM176A TNF TNFRSF1B TNIP1 TPM4 TRAF1 TRIM13 TRPC4 TXNRD1 USP1 WDR92 YBX3 ZNRF1 ZRANB2
- Downregulated:
- ABCC3 ACAA2 C1QA CACNA1A CHD4 COL4A5 CTLA2B CYTIP DAB2 GAB2 GJB1 HCN2 HFE HMGCL HOXA3 IL1R2 ITGA6 LAPTM5 LGALS1 LSP1 MXD4 NFIA NTPCR RHD ROGDI RPL18 SEPT9 UBE2R2
- (Rodríguez et al, 2007) [153]
Macrophages
- Ca2+ -dependent production of ROS and nitric oxide (NO)
- Linked to persistence rather than active replication
- Macrophages and dendritic cells
- APC most likely responsible for priming a Cpn-specific CD8+ CTL response
- Endogenous MHC class I pathway
- Can process and present exogenous Ag to CD8+ T cells
- CD8+ CTL primed by endogenously processed Ags are the most likely to recognize all Cpn-infected cells [120]
Induction of mitogen-activated protein kinases (MAPK) p38
- Extracellular signal
- Regulated kinase ERK1/2 and c-Jun N-terminal kinase JNK
- Consequence of the infection in various cell types
- Ubiquitous stress-related signaling proteins
- Essential step in the promotion of a proinflammatory and often proliferative phenotype [60]
- Participate in different manifestations of the bacterium-induced phenotype changes of the host cell
- Cytokine, chemokine and adhesion molecule expression
- Foam cell formation
- Increased proliferation [60]
- C. pneumoniae entry
- ERK1/2 activation has been reported
- Crucial for successful internalization of EBs
- Reorganization of actin microfilaments
- C. pneumoniae entry to epithelial cells
- Prevented by a small molecule inhibitor of ERK1/2 activation
- By its upstream activator MEK [60]
Mast cells
- Facilitating immune cell infiltration into the airspace
- Providing a more favorable replicative environment for C. pneumoniae [5]
- Recognize bacterial pathogens through TLRs and NOD-like receptors [5]
- Produce proinflammatory cytokines [5]
- Important role in host defense against bacteria [5]
- Tryptase, chymase
- Degrade many proteins
- Extracellular matrix
Chymase (MCPT4)
- Activate matrix metalloprotease (MMP)-9
- By cleaving a specific site of the catalytic domain of MMP-9
- Chymase inhibitors
- Reduce both the pro- and active forms of MMP-9
- Attenuate its enzymatic activity in vivo
MMP-9
- Necessary for
- Neutrophil infiltration into the lung and airspace
- Intratracheal migration of dendritic cells
- Degrades claudins
- Adhesion molecules important in cell–cell tight junctions
- During influenza-induced lung injury
- Several claudin family members are expressed in lungs
- Claudin-5 and -18
- Components of alveolar tight junctions
- Degradation important for paracellular permeability and cell transmigration [5]
- MC degranulation is required for C. pneumoniae–induced immune cell infiltration into the airspace [5]
- C. pneumoniae množení a šíření was reduced significantly in cromolyn-treated mice [5]
- Lead to unproductive inflammation and greater bacterial growth [5]
- Facilitated the influx of neutrophils and other macrophage/monocytes into the airspaces where the bacterial progeny of the first round of replication could infect them [5]
- Mastocyty v plicích + = + plicní propustnost = + imunitních buněk v alveolech a bronších
- Nevýhoda: Infekce které více napadají imunitní buňky než plicní epitel budou díky tomu pravděpodobnější a silnější
- Chlamydia pneumonia
- H1N1 influenza A virus [5]
- Streptococcus pneumonie [5]
- Výhoda: Bakterie a viry které se množí spíše v epitelu budou vyšší imunitní aktivací více potlačeny
- Mycopl. pneumoniae [5]
Mitochondrie a C.p. infekce
- Apoptosis regulation through the mitochondrial pathway in Chlamydia (Sharma and Rudel, 2009)
- Chlamydia trachomatis induces ROS production
- Beneficial for their development (Abdul-Sater et al., 2010; Boncompain et al., 2010; Chumduri et al., 2013) [154]
- C. trachomatis infection preserved the mitochondrial network (Chowdhury et al., 2017) [154]
- Production of ROS in Chlamydia pneumoniae
- mitochondrial dysfunction is implicated with inflammasome activation in C. pneumoniae infected macrophages (Shimada et al., 2011) [154]
- C. pneumoniae infection resulted in a significantly decreased maximal respiration
- Compared to non-infected control cells at 24 and 48 hpi
- Is characteristic for mitochondrial dysfunction
- A significant increase in the mitochondrial TMRE intensity
- Marker of mitochondrial hyperpolarization
- Observed in C. pneumoniae-infected cells compared to non-infected cells at 48 hpi
- Increase in the mitochondrial membrane potential in C. pneumoniae-infected cells
- Associated with an increased ROS formation
- C. pneumoniae infection is accompanied by a mitochondrial dysfunction !!! [154]
- C. pneumoniae infection of mouse macrophages induced mitochondrial dysfunction (Shimada et al., 2011)
- C. pneumoniae infected HEp-2 cells exhibit a mitochondrial dysfunction, characterized by
- Decreased maximal respiration
- Hyperpolarization of the mitochondrial membrane potential
- Enhanced ROS generation [154]
- The decrease in maximal respiration was specific for active chlamydial growth
- Heat-inactivated C. pneumoniae did not affect the maximal respiration [154]
- Inhibition of host ATP synthase
- Increases the mitochondrial membrane potential (Rego et al., 2001)
- Switches ATP production to glycolysis (Brand and Nicholls, 2011) [154]
- Increased energy demand of C. trachomatis-infected cells
- Provided by elongated mitochondria as a result of higher fusion/fission ratio and that mitochondrial ATP is essential for C. trachomatis growth (Chowdhury et al., 2017) [154]
- C. pneumoniae infection
- mitochondrial dysfunction seems to be the preferred phenotypic state
- Different metabolic characteristics might occur due to the different tissue tropism and differences in their genome [154]
Modifying LDL cholesterol
- C. pneumoniae and/or low density lipoprotein in five study models using differential display and real-time RT-PCR. Lim and Chow, 2006
- Modified genes:
- ACTG1 ARPP19 BCL2A1 CDC27 CDC42EP3 CDK1 CTSH FLOT1 GPR6 IK IRS2 PHF20 RPL27A SLAMF8 UBAC1 UBE2T
Monocyty - blokáda apoptózy
- Successfully escape from programmed cell death
- Inhibiting apoptotic signaling cascades within various host cell populations
- Monocytes
- Epithelial cells
- Microglial cells
- Infected monocytes produce:
- Pro-inflammatory :
- IL-1,
- IL-6
- TNF alfa [65, 99]
- Anti-inflammatory:
- IL-10 cytokines
- Immunosupressive regulator of cell-mediated immunity
- T-cell differentiation to Th2 cells but not to Th1 cells [65]
- IL-10 is a powerful inhibitor of IFN-gamma
- X eradikace chlamydií
- X activation of monocytes
- Involved in the resistance of C. pneumoniae-infected human monocytes to apoptosis [65]
- Induce activation of antigen-specific cell-mediated responses
- Promote the eradication of C. pneumoniae infection [65]
- When Cpn-infected THP1 human monocytes were compared to control uninfected monocytes at 48 hours post-infection
- 17 genes were found to have a significant 4-fold or greater expression
- no gene expression was found to be down-regulated.
- Cytokine secretion
- (IL-1ß, IL-6, IL-8) appears to be maintained for an extended period of infection. [154]
Differentially expressed genes in the epididymal AT transcriptome of combined chronic C. pneumoniae-infected mice
- Upregulated
- AVPR1A C7 IGK-C LRRN1 SLC15A2
Neutrofily
- Neutrophil depletion promotes bacterial clearance during C. pneumoniae lung infection [5]
- Neutrophils are a critical site of C. pneumoniae replication [5]
- Neutrophils play an important role as an infectious target of C. pneumoniae [5]
- TREM-1 expression on neutrophils
- Required for transepithelial migration [5]
- In vitro granulocytes
- Internalize Chlamydia in an opsonin-independent manner
- Phagocytosed Cp were not killed [49]
- Ingested bacteria survived and multiplied within PMN
- Uninfected granulocytes became apoptotic within 10 h [49]
- Infected PMN survived up to 90 h [49]
- Coincubation with Cp significantly decreased the ratio of apoptotic PMN [49]
Antiapoptotic effect of C.pneumoniae infection was associated with:
- Lower level of procaspase-3 processing [49]
- Reduced caspase-3 activity in infected PMN
- LPS major component in antiapoptotic effect [49]
- “Trojan horses” for subsequent infection of other cells, such as macrophages [49]
- Life span of mature neutrophils can be extended in vitro by
- Incubation proinflammatory cytokines
- GM-CSF
- G-CSF
- IL-8
- IL-1ß [49]
- Glucocorticoids
- Bacterial products
- LPS
- FMLP [49]
- C. pneumoniae activates
- P38 MAP kinases
- PI3K/Akt and ERK1/2
- Essential to initiate the apoptosis delay in C. pneumoniae-infected neutrophils
- Involved in the maintained expression of the anti-apoptotic protein Mcl-1
- PI3K/Akt pathway
- Activation of NF-?B-dependent release of IL-8 by infected neutrophils [152]
Další mikrobi a apoptoza neutrofilů
- Several microbial pathogens reported to influence cellular apoptosis
- Indukují apoptozu:
- Escherichia coli
- Candida albicans
- Inhibition of apoptosis by intracellular pathogens
- HGE
- L. major
- Chlamydias etc. [49]
Host cell responses to Chlamydia pneumoniae in gamma interferon-induced persistence
- Overlap those of productive infection
- Are linked to genes involved in apoptosis, cell cycle, and metabolism (Eickhoff et al, 2007) [153]
- Regulated at various time points:
- ASNS BNIP3 CA9 CTGF CTH CXCL11 CYR61 DACT1 DDX58 DEPTOR DKK1 EGR1 ETS2 FOSL1 FST FZD10 HERC5 HERPUD1 HEY1 HSPA1A HSPA1B IFI44 IFIT4 IL11 INSIG1 ISG15 ISG20 KRT17 LOX LYPD3 METTL7A MXRA5 NDRG1 NFIL3 OAS1 OASL PDLIM5 PHLDA1 PLK2 PPP1R3C PSPH PTGER4 PTPN13 RAI3 SLC1A4 SREBF1 STC2 TNFAIP3 TRIB3 VEGFA ZC3HAV1
Interakce s organelami
- Indicate that interaction for optimal chlamydial replication required with
- Multivesicular bodies,
- Golgi-apparatus,
- Mitochondria,
- Lipid droplets
- Membrane fusion events between host-derived vesicles and the inclusion
- May deliver nutrients to Chlamydia. [173]
Golgiho aparát
- Golgin-84 is proteolytically processed during infection through activation of
- Host caspases calpains
- Potentially by bacterial protease(s)
- Leads to Golgi fragmentation into ribbon-like structures
- In a process mediated by Rab-6 and Rab-11
- These Golgi ministacks are recruited around the inclusion
- Formation of Golgi-ministacks is necessary for chlamydial uptake of host sphingolipids and replication
- Golgi-derived exocytic vesicles containing sphingolipids and cholesterol fuse with the inclusion. [173]
Oxidační stres
- ROS play a central role in the induction of the pathological consequences of C. pneumoniae infection
- C. pneumoniae infection increase ROS in various cell types:
- Bronchial epithelium
- Platelets
- Macrophages [60]
- C. pneumoniae infection is known to induce an increase in host cell ROS levels
p38 MAP kinases activation
p47 GTPases
- Regulators of immunity to intracellular pathogens
- Regulate innate immunity and inflammation during murine chlamydial infection (Nelson et al., 2005; Bernstein-Hanley et al., 2006; Miyairi et al., 2007) [132]
Germline-encoded pattern recognition receptors (PRRs)
- Detect pathogen-associated molecular patterns
- Signal the presence of a foreign microorganism to the host TLRs [6]
- Best-characterized class of PRRs [6]
Stopové prvky
- In the patients’ valves, iron, magnesium, and zinc
- Each correlated to calcium
- A marker of the histopathological severity of ateroslerotic disease
- Patients showed
- 10- to 70-fold increases of these trace elements in valves
- Increased copper/zinc ratio in serum
- All patients had
- Disturbed trace element balance in valves and serum
- Suggestive of active immune process and infection
- The 20-fold increase in iron, essential for C. pneumoniae growth, in sclerotic valves
- Suggests a new possible link to this infection in aortic sclerosis [169]
Systemic dissemination
- Mediated by peripheral blood mononuclear cells (PBMCs)
- Alveolar macrophages
- Infected by granulocytes
- Migrate through the mucosal barrier
- Using lymphatic tissue
- Access to the systemic circulation as PBMCs
- Reaching the spleen and the vasculature [25]
- Excellent capacity of C. pneumoniae to survive in different immune cells
- Use the immune cells as carriers for breaching the blood-tissue barriers
- Infected monocytes transmit the pathogen
- Endothelial
- Smooth muscle cells [49]
- Cpn can infect and multiply in nonprofessional APC, including
- Airway epithelial
- Endothelial
- Smooth muscle cells [120]
- Chlamydie lze detekovat v granulocytech, alveolárních makrofázích a alveolárních epiteleliích
- Déle než 8 týdnů po infekci
- Infikované makrofágy procházejí slizničními bariérami
- Otevírají mikrobu lymfatický systém
- Infikované periferní monocyty
- Vektorem infekce i uvnitř krevního řečiště - do stěny cév a dalších orgánů [121]
TLR2
- Chlamydia pneumoniae infection promotes vascular smooth muscle cell migration through a Toll-like receptor 2-related signaling pathway (Wang et al, 2013) [153]
- Upregulated:
- BAIAP2 CCL20 CX3CL1 GREM2 ID1 ID2 MCM6 MCM7 NQO1 PCNA TLR2
- Downregulated:
- AKAP12 ANXA1 CD55 DUSP1 EDN1 EGR1 ERRFI1 F3 FOS GJB2 LOC683626 NFIL3 PKIG PPAP2B PSAT1 RASL11B STMN2 TGFA TGFB3 THBD
Proatherogenic pathogens on adipose tissue transcriptome and fatty acid distribution in apolipoprotein E-deficient mice
- (Hyvärinen et al, 2013) [153]
- Upregulated
- C230075M21RIK EIF2AK2 FCRLS GPC3 IFIT3 OSR2 PPAP2B USP18
- Downregulated
- 8430408G22RIK ACTR3 DDIT4 GM14121 Gm5277 Gm9143 GPX8 IGKV4-54 LOC665281 MOD1 RPS3A YWHAG
Železo a infekce C.p. myší
- Transferrin receptor (TfR), Hepcidin (Hamp1), and ferroportin 1 (Fpn1)
- Increased in the liver on day 4
- Normalized except for TfR that tended to decrease
- TfR responses were similar to Fe in serum
- Increased on day 4
- Tended to decrease thereafter
- In the liver
- Fe was increased on day 4 and also on day 40
- Reappearing increases in Cu/Zn on day 40
- Concomitant with the increase in liver Fe on day 40
- TfR tended to decrease
- Viable C. pneumoniae was present in the lungs and liver
- May indicate the early phase of activation of recurrent infection [175]
- The circulating peptide hormone hepcidin
- Produced by the liver
- Regulates the ferroportin 1 (Fpn1) levels
- Transferrin receptor (TfR)
- Marker of the Fe status
- Regulates the ferroportin 1 (Fpn1) levels
- Infection and inflammation
- Liver hepcidin levels in mice increase during acute C. pneumoniae infection
- Associated with concomitantly altered Fe levels
- High Fe levels may not only predispose to C. pneumonia infection
- Could enhance the effects of C. pneumonia on atherogenesis [175]