nemoci-sympt/BAKTERIALNI-INFEKCE/chlamydie/stavba-chlamydia-pn
Druhy
{kind=link}
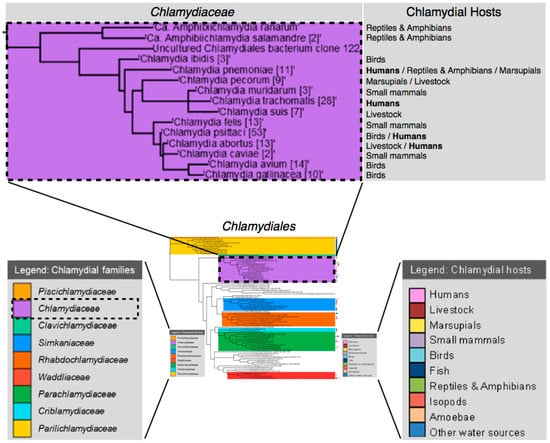
- Chlamydiaceae has two genera
- Chlamydia
- Chlamydia trachomatis
- Chlamydophila
- Chlamydia pneumoniae
- Non-Chlamydiaceae
- Simkaniaceae
- Waddliaceae
- Parachlamydiaceae
- Chlamydia pneumonia - (PN)
- Chlamydophila pneumoniae: human, koalas, horses, lower vertebrates [99]
- Chlamydia pneumoniae - new syn. Chlamydophila pneumoniae
- C. pneumoniae, previously known as C. psittaci (strain TWAR) [22]
- Chlamydophila psittaci : birds, sheep, cattle, cats [99]
- Chlamydia trachomatis - (TR)
- Chl. suis - (SU)
- Chlamydophila caviae
- Guinea pigs inclusion conjunctivitis [11]
- Chlamydophila abortus
- Abortions and fetal loss in ruminants
- Spontaneous abortions and respiratory disease in humans [11]
- Chlamydophila felis
- Pneumonia and conjunctivitis in cats [11]
Phylogenetic studies
- Chlamydia likely shares a common ancestor with cyanobacteria
- Group containing the endosymbiont ancestor to the chloroplasts of modern plants
- Chlamydia retains unusual plant-like traits
- Genetically and physiologically [137]
- Enzyme L,L-diaminopimelate aminotransferase
- Related to lysine production in plants
- Linked with the construction of chlamydial cell walls [137]
- Genetic encoding for the enzymes remarkably similar in
- Plants
- Cyanobacteria
- Chlamydia [137]
- Close common ancestry [137]
Genetika
- Chl. pneumonia shares less than 10% of the DNA homology with Chl. psitaci, trachomatis [3]
- Highly reduced genome of approximately 1 Mb
- Inability to introduce gene-specific DNA [1]
- High degree of genetic conservation
- Very limited evidence of horizontally acquired foreign DNA [1]
- Chlamydiae are highly recombinogenic [1]
- Recombination occurs naturally
- Clinical resistance might spread rapidly following an initial introduction of an exogenous resistance gene into the chlamydial population [1]
- From 167 genes
- 263 C. pneumoniae proteins encoded
- Identified by matrix-assisted laser desorption ionization-mass spectrometry (MALDI-MS) [21]
- Replicate by binary fission
- Contain DNA, RNA, and ribosomes
Horizontal gene transfer (HGT)
- Exact mechanism of HGT of Chlamydia is largely unknown
- HGT and subsequent recombination can occur after co-infection within host body
- Discovery of a number of phages has shown that phage-mediated transduction in Chlamydia is possible as well
- Chlamydia genomes contain full gene sets necessary for HGT
- Numerous feasible HGT events that has occurred in evolutionary lineages of Chlamydia [143]
Hotspot for genome variation - “Plasticity zone”
- Encodes virulence factors
- Membrane attack complex/perforin protein (MACPF)
- Cytotoxin
- Genes related to important biosynthesis and salvage pathways [143]
- Presence or absence of these genes in the region
- Confer different niche-specificity to the organisms [143]
- Chlamydia interact frequently with membranes during their infection
- MACPF is important
- Gene transfer of MACPF from C. abortus S26/3 to C. felis Fe/C-56 (SV: 1.0) and to C.psittaci M56 (SV: 1.0) separately [143]
- tryptophan biosynthesis pathway
- Necessary for survival of Chlamydia since host restricts chlamydial growth by degrading tryptophan as a defense mechanism
- Each Chlamydia species possesses different level of functional gene sets of tryptophan biosynthesis pathway
- TyrP gene encoding tyrosine/tryptophan transport protein was transferred within Chlamydia genus (from C. felis Fe/C-56 to C. caviae GPIC; SV: 1.0)
- TrpC of C. pecorum E58, C. felis Fe/C-56, and C. caviae GPIC were might have been transferred from Coxiella burnetti dugway 5J108-111 strain
- TrpC encodes indole-3-glycerol phosphate synthase
- Necessary in the fourth step of tryptophan biosynthesis pathway [143]
- Adenosine deaminase (Add) associated in purine ribonucleotide biosynthesis pathway
- Tox/adhesion may affect Chlamydia pathogenesis and host-range
- Occurred from Escherichia coli and Citrobacter rodentinum to C. caviae GPIC (SV: 1.0).
- T3SS genes seemed to be mostly transferred within Chlamydia
- Structural components of the T3SS apparatus
- Intra-genus HGT in 5 genes (SctW, SctS, SctR, SctF, and SctP) [143]
- Effector proteins for manipulation of host cell immune response
- EEA1, Cap1, CPAF, Tsp, and pGP6-D
- Appear to have histories of HGT only with other Chlamydia [143]
- Exchanges of virulence related genes
- Mainly occurred within Chlamydia genus
- Intra-genus HGT may have been a major mechanism
- Transfer of those genes may provide a fitness benefit to the recipient [143]
Elementární tělíska - “elementary body” - EB
- Infekční forma - extracelulární
- Metabolically inactive [7]
- Rigid cell wall
- Disulfide cross-linking of envelope proteins [23]
- Survival outside of the host cell
- Bacterial nucleoid is highly compacted
- Condensation of nuclear material [52]
- By the bacterial histone-like proteins HctA and HctB
- Nucleoid has an eccentric location in the cell body
- Association with the bacterial inner-membrane or cell wall [52]
- Little or no peptidoglycan is present in the cell wall
- Structural rigidity due to the highly cross-linked nature of the outer-membrane complex
- Inter and intramolecular cystine bonds
- Between the cysteine rich proteins of the outer envelope:
- OmpA, OmcB, and OmcA [52]
- Hexagonally arrayed protein layer
- Predominantly OmcB
- At the inner surface of the outer-membrane complex
- Contribute to cellular stability in EBs [52]
- Extracellular EB form of the bacterium is known to be sensitive to reducing agents (Kim et al., 2009; Di Pietro et al., 2013)
- Envelope structure varies between the different life cycle forms of Chlamydiae (Hatch, 1996)
Buněčná stěna EB
- Obaleny dvojitou membránou - znakem gramnegativních bakterií
- Nemají však mezi oběma membránami vrstvu peptidoglykanu
- Nemají proto pevnou buněčnou stěnu [121]
- (EBs) consists of densely cross-linked cysteine-rich proteins (CRPs)
- Might respond to the osmotic stability of EBs
- Present only in EBs but absent in dividing RBs (Hatch et al., 1984):
- Cell walls with inner and outer membranes [25]
- Many similarities with other Gram-negative bacteria [1]
Peptidoglykan
- Possess nearly the complete set of genes required for the synthesis of peptidoglycan
- Jen stopová množství muramic acid a jen někt. studie [100]
- Needed for RB division [100]
- Chlamydia lacks the peptidoglycan (PG) in their cell wall
- Major outer membrane protein (MOMP) may substitute to the PG (Garrett et al., 1974; Newhall & Jones, 1983; Hatch et al., 1984)
- There is functional PG in the cell wall of Chlamydia trachomatis (Liechti et al., 2014)
- V genomové sekvenci C. trachomatis sekvence genů pro syntézu peptidoglykanu objevena byla
- Může vysvětlit, že vývoj chlamydií je in vitro blokován betalaktamovými antibiotiky likvidujícími bakterie s peptidoglykanem v buněčné stěně
Penicilin vázající proteiny (PBPs)
- Ve stěně chlamydií byly prokázány
- Předpokládá se i přítomnost proteinové sítě podobné peptidoglykanové výztuži
- Peptidoglykan by mohl být potřebný pro dělení RTs [121]
Inclusion membrane surrounding C. pneumoniae
- Additional barrier towards antibiotics
- Does not allow entry of componentsof size of greater than 500 Da by passive diffusion [60]
- Small size
- Tissue penetration
- Ability to pass through eukaryotic cell membranes
- Necessary for any therapeutic agent targeting structures of replicating Chlamydia spp. [60]
Chlamydial outer membrane
- Family-specific lipooligosaccharides
- Permeability barrier limits the entry of antibiotics into gram-negative bacteria
- Zevní mebrána obsahuje proteiny označované jako proteiny zevní membrány (outer membrane proteins, Omps)
- protein containing an RGD sequence and also were classified as outer membrane proteins
- Made of cysteine-rich proteins
- Form a network of both intra and inter molecular disulfide links
- Contributes to the integrity [151]
- Be potential vaccine candidates
- Defined as proteins that had more than 3.0% cysteine in their primary amino acid sequence, above the mean genomic ORF cysteine content
- Genové sekvence: ORF 1290, ORF 1294, ORF 1296 [151]
- Small proteins that have cysteines in their N- and C-terminus
- May contribute to the network formed by disulfide linkages
- May be anchored in the outer membrane via their N-terminus
- May have their C-terminus exposed
- Can interact with the host cells [151]
Major outer membrane protein - MOMP
- Porin channel in the outer membrane of Chlamydia species [125]
- Nejhojněji zastoupen velký protein zevní membrány
- Obsahuje druhově a poddruhově specifické epitopy
- Může být identifikován monoklonálními protilátkami
- MOMP C. pneumoniae méně imunogenně a antigenně komplexní než MOMP ostatních chlamydií
- Reduced levels of MOMP
- Could enable chlamydiae to avoid the development of protective immunity [39]
- Within 1 h of infection, the MOMP in the EB cell wall is reduced to monomers [99]
- Synthesised late in the developmental cycle
- Present only in EBs
- Disulfide bonds and MOMP
- Maintain the structural integrity of EBs lacking the peptidoglycan
- Most abundant protein in the outer membrane of chlamydiae
- Form pore-like structures
- Depending on the disulfide bonding [100]
- Major immunodominant antigen
- Four variable domains
- Basis for serotype differentiation [100]
- Highly conserved
- Neutralising antibodies
- Produced against surface-exposed C. pneumoniae MOMP epitopes
- Epitopes are conformational and easily destroyed by detergents [100]
- Maintaining cell wall rigidity
- Consistence of MOMP varies between the different life cycle forms of the bacteria
- MOMP of EBs
- Tightly cross-linked with disulfide bridges
- Insoluble in sodium dodecyl-sulfate (SDS)
- Detergent used to solubilize chlamydial inner membrane proteins in the absence of mercaptoethanol (Hatch et al., 1981)
- Solubility of MOMP in the presence of mercaptoethanol
- Remarkable sequence similarity between the MOMPs of chlamydial species
- C. pneumoniae MOMP does not seem to be as immunodominant as Chlamydia trachomatis MOMP [124]
- 40 kDa
- MOMP elicits antibodies that recognize both linear and conformational antigenic determinants
- Not surface exposed and is immunorecessive
Omp2 - OmcB/EnvB
- Později ve vývojovém cyklu jsou syntetizovány na cystein bohaté Omp2 a Omp3 [121]
- cystein-rich outer membrane proteins (Omp)
- protein zevní membrány
- Ochrana proti zevnímu prostředí
- 62 kDa
- Identified as a target of immune recognition in both C. trachomatis and C. pneumoniae infections [124]
Omp3 - OmcA/EnvA
- Později ve vývojovém cyklu jsou syntetizovány na cystein bohaté Omp2 a Omp3 [121]
- cystein-rich outer membrane proteins (Omp)
- Disulfidové crosslinking vazby
- protein zevní membrány
- Ochrana proti zevnímu prostředí [121]
LPS - Chlamydial LPS (cLPS)
- Main antigenic component of Chlamydiae (Nurminen et al., 1983)
- CLPS is less immunogenic than other bacterial LPS (Kalayoglu, et al., 2000).
- Induces the autocrine production of the antiapoptotic cytokine IL-8 [49]
- Produlžuje život neutrofilů - hostitelské bu. [49]
- Following azithromycin exposure
- Residual chlamydial envelopes can persist in inclusions for up to 4 weeks
- Metabolically active reticulate bodies are effectively destroyed [35]
- Presence of chlamydial LPS could provide a source for sustained inflammation
- Chemotaxis of polymorphonuclear leukocytes is stimulated by epithelial cells containing residual envelopes [35]
- Purified LPS was shown to induce tumor necrosis factor alpha production from whole blood [35]
- In both EBs and RBs [100]
- Chlamydial LPS is a far less potent endotoxin and inducer of an acute immune response
- Accumulate in the plasma membrane of infected cells (Karimi et al., 1989)
- Decreased membrane fluidities
- Endocytic processes
- Lysosome-endosome fusion and complementmediated cytolysis
- Bacteria’s way of hiding from the host cell’s immunity system
- Chlamydial LPS evokes a weaker immune response (Kalayoglu, et al., 2000)
- Hlavním endotoxinem gramnegativních bakterií je lipopolysacharid (LPS) lokalizovaný na povrchu chlamydií
- Jak u ET, tak u RT
- Strukturálně podobný formám LPS enterobakterií
- Společné, tak rodově specifické epitopy
- Mnohem nižší aktivitu [121]
LPS rough form
LPS smooth form
Polymorphic membrane proteins (Pmps)
- 9 Pmp genes in C. trachomatis
- 21 genes in C. pneumoniae [100]
- Some Pmps are potentially surface-exposed and immunogenic
- V zevní membráně
- Každý ze 3 studovaných druhů chlamydií má rodinu vzdáleně příbuzných genů pro Pmps
- Rodina Pmps genů sestává z heterogenní skupiny genů
- S nízkou identitou
- S podobnými charakteristikami
- Mnohé z genů kódují proteiny o velikosti 90–100 kDa [121]
GroEL1
- Localized on the surface of EBs
- Adheze na epiteliální buňky ? [13]
- Preincubation of HEp-2 cells with rGroEL1
- Significantly reduced subsequent infection with C. pneumoniae
- Adhesion of infectious bacteria to eukaryotic cells was not affected [13]
Heat shock protein 60 - cHSP60
- 60 kDa
- Activates Toll-like receptor 4 (TLR4)
- Signalizace via MyD88 pathway in vitro
- Stimulates the production of pro-inflammatory cytokines in epithelial cells
- CHSP60 induced IL-6 release and CD80 and CD86 expression in WT cells
- CHSP60 promotes lung inflammation and DC activation via TLR4 and MyD88 [9]
- Induce DC maturation [13]
- Synthesis and release of chlamydial hsp60 from
- Persistently or repeatedly infected mucosal epithelial cells
- Alveolar macrophages
- Provide a prolonged antigenic stimulation
- Strongly amplifies chronic inflammation
- Leads to tissue damage and scarring in the asthmatic lungs [22]
- C. pneumoniae gene products (mainly heat shock protein-60)
- Through the activation of TF (notably NKF kappa-B)
- Activation of most cellular elements in bronchial tissue
- Cascade of cytokine release and adhesion molecule upregulation
- Favours cellular influx into the airways, persistent infection and airway remodelling [25]
- Brání apoptóze
- Key antigen in chronic chlamydial infections
- Scarring tissue damage in trachoma is associated with Hsp60 targeted immune responses [99]
- Localizes in human atheromatous tissue [99]
- Associated with the development of atherosclerosis [99]
- Persistent infection model affected by interferon gamma:
- Hsp60 kept its production near a normal level [99]
- Synthesis of the MOMPs, Omps, lipopolysaccharides and other structure proteins were greatly reduced [99]
- C. pneumoniae HSP60, a protein produced by reticulate bodies in chronic infection
- Contribute to LDL oxidation and endothelial dysfunction
- Through oxidative stress
- Stimulate VSMC proliferation and inflammatory responses through Toll like receptor activation
- Target for prevention of CVDs associated to C. pneumoniae-induced oxidative stress [105]
- proteiny teplotního šoku (heat shock proteins, Hsps)
- Zajišťující univerzální stresovou odpověď buňky
- Naklonovány geny kódující proteiny Hsp10, Hsp60 a Hsp70
- Všechny tři Hsps lze nalézt v komplexu zevní membrány ET i RT
- Hsp60 a Hsp70 jsou v průběhu přirozené infekce vysoce imunogenní [120]
Brání apoptóze infikovaných buněk
- Heatshock protein 60
- Blockade of mitochondrial cytochrome c release and caspase 3 activation [55]
- Critical for bacterial survival [55]
- Activate anti-apoptotic proteins such as:
- Bcl-2
- NF-kappa B
- Expression of multiple genes involved in inflammatory responses [56]
- Genes in anti-apoptotic mechanisms [56]
- Chlamydia-infected host cells are resistant to pro-apoptotic stimuli such as
- TNFalpha
- Fas antibody
- Staurosporine
- UV-light [56]
Elevated antibody levels to chlamydial hsp60
- Associated
- Scarring trachoma
- Pelvic inflammatory disease (PID)
- Tubal factor infertility
- V.s. pathogenesis of atherosclerosis [22]
- Tissue damage follow both recurrent and persistent infections
- Largely the consequence of the immune response to chlamydial hsp60
- Production of proinflammatory cytokines released by the infected cells
- TNF-alfa
- IL-1beta
- IL-6
- IFN-gamma
- Liberation of cellular constituents from lysed cells
- Cross-reactivity with HSP60 from either the human host or other pathogens can happen [39]
12 kDa protein
Glycolipids
Phospholipids
Fatty acids
Vnitřní membrána
- Structure of the cytoplasmic membranes and of the wall of bacteria is dependent on the associated proteins
- Impermeable to
- water
- water-soluble substances
- Small-sized molecules
- Ions, small inorganic molecules
- Peptides or proteins
- To enter into or to interfere with a cell or a bacterium
- A ligand - receptor interaction mus happen
- Small molecules such as sugars
- Small peptides
- Antibiotics
- Heavy metals
- Double lipid layer structure of the membrane
Infikované buňky
- Epithelial and endothelial cells
- Macrophages / monocytes [12, 162]
- Dendritic cells (DCs) [12]
- Mucosal epithelial cells
- Endothelial
- Smooth muscle cells
- Cardiac muscle cells [48]
- Alveolar epithelial cells type II
- Major target cells for C. pneumoniae in chronic but not in acute respiratory infection
- In COPD patients [159]
- Human coronary artery endothelial cells [162]
- Smooth muscle cells [162]
- Human embryonic fibroblasts [162]
- Disseminates to and can be metabolically active in the liver
- Presence of C. pneumoniae in the liver may influence the metabolism and the trace element balance in the body
- C. pneumoniae DNA was demonstrated in the liver even on day 40. [175]
Adheze na buňku
- Pivotal step in pathogenesis
- Extreme high and low temperatures can reduce the attachment [99]
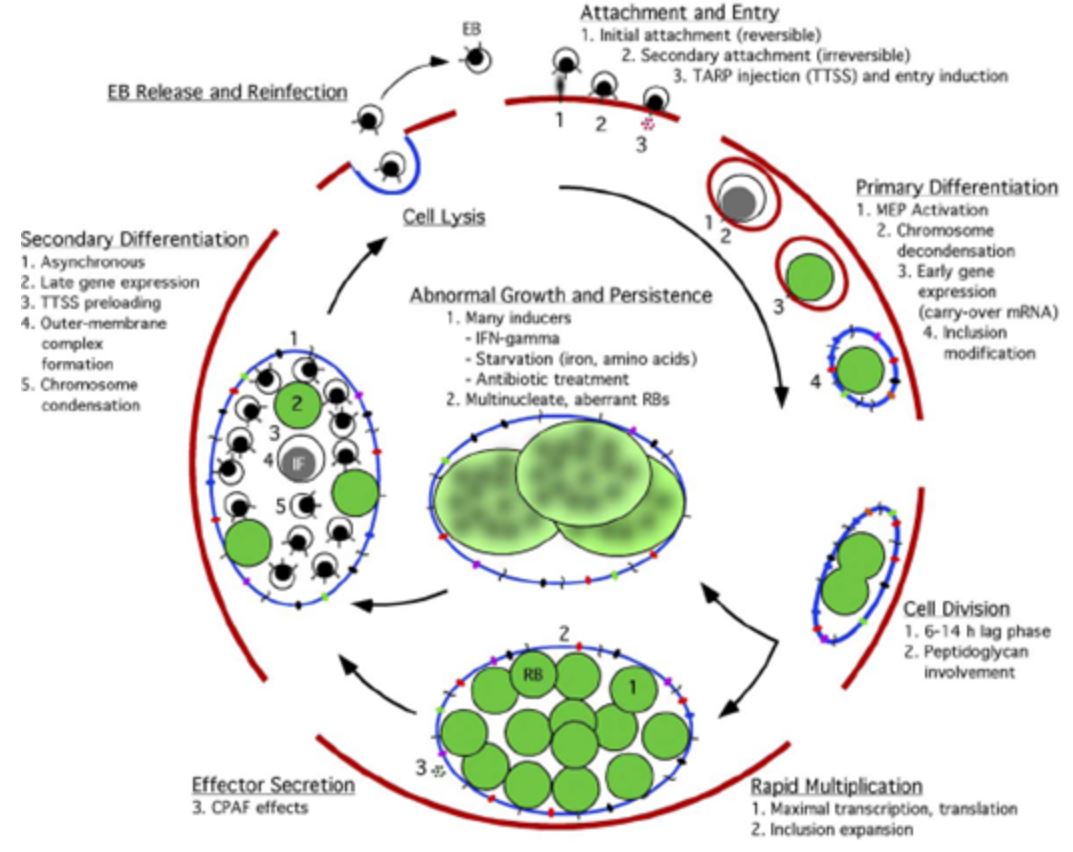
1. Initial attachment of chlamydiae to cells
- Reversible electrostatic interactions with heparan sulfate-like glycosaminoglycans
- Electrostatic interactions of the bacteria with heparan sulfate containing glycosaminoglycans
- Reversible
- Chlamydia bacteria use glycosaminoglycans (GAGs) as receptors for cell attachment (Wupperman et al., 2001)
- Ubiquitously on the surface of eukaryotic cells
- Heparan sulfate-like GAG spec. pro Chlamydii
- Occur through binding to the surface exposed OmpA protein [52]
OmcB protein chlamydiální
- Adhesion of the infectious elementary body to human HEp-2 cells
- interacting with heparin/heparan sulfate-like glycosaminoglycans (GAGs)
- Basic amino acids located in the first of a pair of XBBXBX heparin-binding motifs
- Position 57 (arginine) in the first XBBXBX motif
- Position 61 (arginine) in the second motif
- lysine 69 C terminal - key roles in the interaction [6]
- Discrimination between heparin-dependent and -independent adhesion
- C. pneumoniae OmcB apparently interacts with domains of heparan sulfate harboring a diverse subset of O-sulfations [6]
Heparan
- Na buňce
Mg2+, Ca2+
- Na buňce
2. Binding to cellular receptors
- Irreversible binding stage
- Chemically mutagenized cell lines
- C. pneumoniae then binds to cellular receptors
- Cell adhesion occurring at 5 min postinfection
Pmp
- Adhesin family of polymorphic membrane proteins (Pmp) in Chlamydia pneumoniae consists of 21 members
- Pmp21 binds to the epidermal growth factor receptor (EGFR)
- Pmps contain large numbers of FXXN
- X is any amino acid
- GGA - I/L/V motifs
- Two of these motifs are crucial for adhesion by certain Pmp21 fragments [3]
- Form of elongated protofibrils
- Thioflavin T fluorescence, like the amyloid protein fragment ß42
- Mutant version of Pmp21-D (D-Mt)
- FXXN motifs replaced by SXXV
- Markedly reduced capacity to form oligomers [3]
- Monomers of both variants exist predominantly as random coils
- Oligomers form predominantly beta-sheets
- Significantly enhanced binding to human epithelial cells
- binds EGFR more efficiently than D-Wt monomers [3]
C. pneumoniae entry - interaction with cell surface receptors
- A total of six membrane proteins overexpressed in skeletal muscle [GEM] play a key role during C. pneumoniae entry
- Chemokine C-X-C motif receptor 7 [CXCR7]
- Integrin beta 2 [ITGB2]
- Platelet-derived growth factor beta polypeptide [PDGFB]
- Vascular endothelial growth factor [VEGF]
- Vascular cell adhesion molecule 1 [VCAM1]
- GTP binding protein [155]
- None alone is essential to prevent entry
- Combination knockdown of three genes significantly inhibits C. pneumoniae entry
- Coding for CXCR7, ITGB2, and PDGFB
- But the entire network is resistant to the six-gene depletion
- Indicating a resilient network [155]
- C. trachomatis serovar E
- Closely associated with protein disulfide isomerase (PDI)
- A component of the estrogen receptor complex [52]
- Attachment dramatically enhanced in estrogen dominant primary human endometrial epithelial cells
- At the apical surface of cells - lumen [52]
- Disulfide isomerase activity of PDI
- May play a role in the reduction of the highly disulfide cross-linked EB OMC
- A requirement for productive attachment and entry [52]
- (M6P)/insulin-like growth factor 2 (IGF2) receptor [7]
- Cellular receptors critical for entry of the infectious elementary body (EB)
- Epithelial membrane protein 2
- Mannose 6-phosphate/insulin-like growth factor 2 receptor
- Platelet-derived growth factor receptor
- Tom complex [155]
3. Signal transduction
- Receptor and actin activity at 25 min
Endocytosis or pinocytosis and Cytoskeleton remodeling
- Endocytosis at 2 h post infection in vitro
- Host cell receptor-mediated [25]
- Enter nonphagocytic cells
- Dependent on
- Temperature
- Functions of microfilaments and microtubules [99]
- Etc.
- Substances inhibiting the functions of microfilaments and microtubules
- Can inhibit the endocytosis of attached chlamydiae [99]
- Tarp (Translocated actin-recruiting phosphoprotein, CT456)
- Rychle vyloučen do cytoplazmy buňky
- Rapidly phosphorylated at tyrosine residues
- Phosphorylation correlates spatially and temporally with actin recruitment
- Recruits actin
- On the cytoplasmic face of the cell membrane while the EB is still extracellular
- V.s. EBs have a functional TTSS that delivers important signaling molecules to the host cell prior to differentiation
- Expression of important proteins must be part of the secondary differentiation process in which RBs are converted to EBs:
- For attachment and entry
- For assembly into functional complexes [52]
- Different steps of actin reorganization during the internalization of chlamydiae
- Cytoskeleton remodeling during endocytosis - 2 family of proteins:
- ADP-19 ribosylation factor 6
- Rho GTPases
- Downstream kinases following these two families of proteins include
- Phosphatidylinositol 3-kinase
- MEK-ERK kinases
- ERK 1/2
- Adaptor protein Shc [155]
- Two other families of proteins are also activated during endocytosis of C. pneumoniae entry
- Focal adhesion kinase (FAK)
- Dynamin
- Bind to several proteins
- Interacting directly or indirectly with F-actin
- Connect the endocytic machinery to the actin cytoskeleton [155]
Macrophage infectivity potentiators (MIPs) - CpMIP
- Virulence factors in intracellular pathogens
- Surface-exposed immunodominant proteins
- On both EBs and RBs
- MIP homologue Cpn0661 is a secreted effector protein
- Providing the MIP protein with access into the host cell cytoplasm
- On the surface of inclusions from 20 to 72 hours post-infection [99]
Bacterial adhesin candidates
- MOMP is suggested to have a role as an adhesin
- OmpA
- OmcB
- Omp2
- Only on chlamydial EBs, is believed to be involved in the attachment, entrance and avoidance of lysosomal fusion [99]
- Hsp70 [52]
- Polymorphic membrane protein (Pmp) family
- autotransporters [52]
- 21 members in C. pneumoniae
- PmpD of C. pneumoniae
- As an autotransporter
- antisera raised to the N-terminal passenger domain blocked chlamydial infectivity [52]
- E.g. heparin sulphate-like proteoglycans
- Glycosylated MOMP
Inkluze
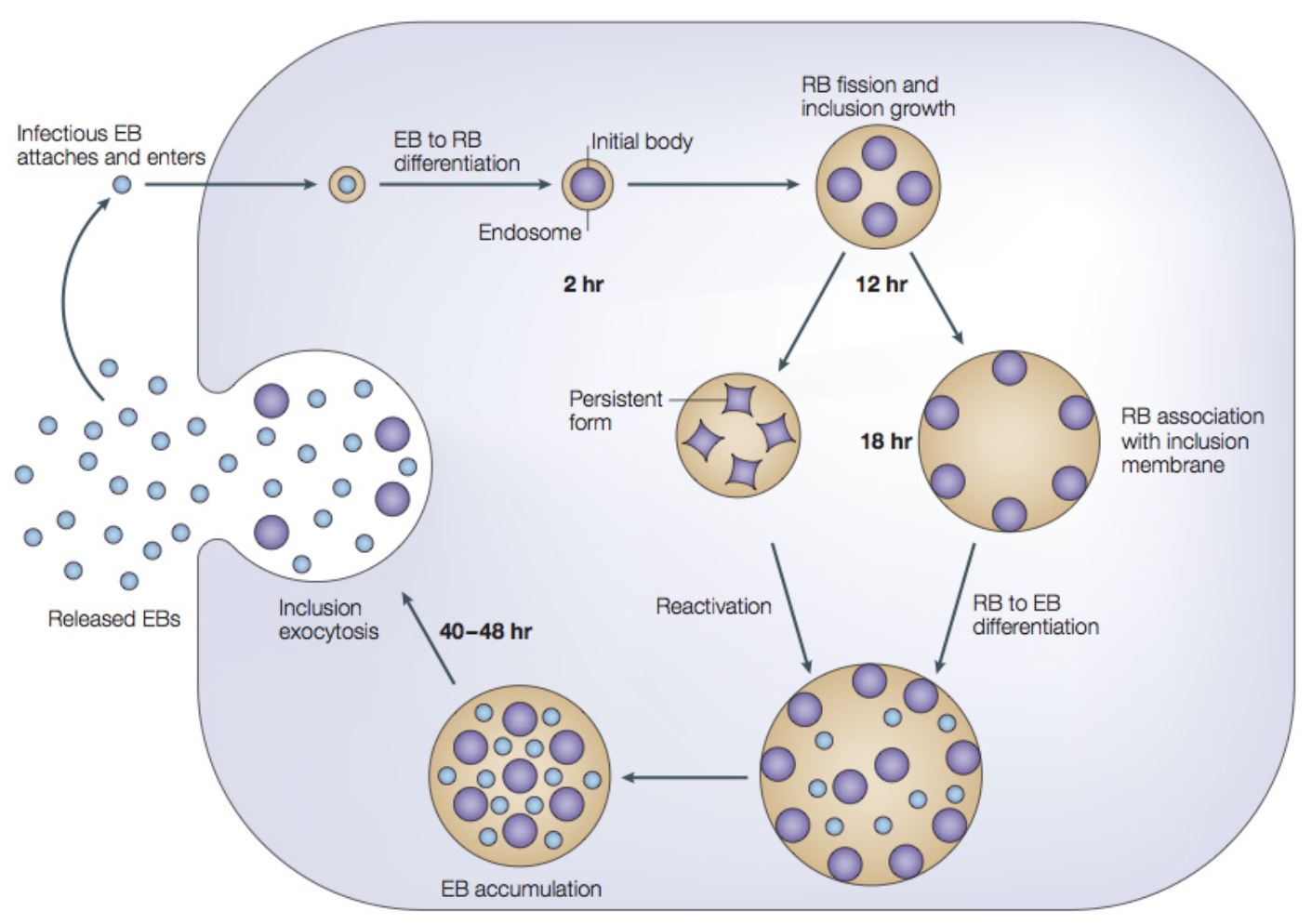
- During the first two hours the EBs internalize into the host cell
- After internalization they remain within individual, tightly membrane-bound vesicles.
- EBs are wrapped in a membrane-bound vacuole to form inclusion
- Enclosed endosomes
- Umí inhibovat endozomální acidifikaci [121]
- Zůstávají ukryty v endozomu - ve vakuole, kterou do buňky vstoupily
- V dalším vývoji se mění v inkluzi [121]
- Avoid phagolysosome formation and thus, digestion of the EBs be prevented [99]
- Escape from the fusion of lysosomes [99]
- Initiation of the differentiation of EBs into RBs take place almost synchronously [99]
- Development occurs within a membrane bound vacuole - inclusion
- Avoid fusion with host-cell lysosomes !!! [34]
- Does not fuse or interact with endosomes or lysosomes during productive growth
- Local aggregation
- Probably associated with the fusion of inclusions [99]
- Microcolony of RB
- Vše potřebuje intracellular ionized calcium ([Ca2+]i) [99]
- Disrupts F-actin/ß-tubulin cytoskeletal association with NF-?B/I?B?
- Inducing a NF-kappa B activation [99]
Stavba inkluze
- Inkluze má plášťový cytoskelet vytvořený sítí F-aktinu a intermediárních filament
- Kooperativě stabilizují vakuolu obsahující patogen
- Chlamydie v.s. kooptují funkci obou vláken ve snaze
- Stabilizovat inkluzi
- Minimalizovat expozici jejího obsahu vrozeným obranným imunitním procesům [120]
- proteiny inkluzní membrány (inclusion membrane proteins, Incs
- První z nich, prokázaný v roce 1995 u C. psittaci, byl nazván IncA
- Od té doby byly objeveny další [121]
- V genomu C. trachomatis bylo odhaleno 46 kandidátů jako potenciálních členů Inc proteinů
- Šest z těchto genů bylo vybráno pro produkci protilátek
- Pět je lokalizováno v inkluzní membráně
- Genom C. pneumoniae obsahuje dokonce vyšší počet hypotetických Inc proteinů
Funkce inkluzní membrány
- Transport vezikul
- Vývoj inkluze
- Obranu proti fúzi s lysozomem
- Získávání potravy
- Signalizaci spojenou s reorganizací ET - RT - ET [121]
Type III secretion system (TTSS)
- Likely way for delivery of several chlamydial proteins into the host cell cytoplasm (Subtil et al., 2005)
- Secreted proteins or even components of the TTSS
- Can be considered as vaccine candidates
- Potential CD8 T-cell antigens of particular interest in vaccine development
- CPAF
- LcrE (CopN) (Sambri et al., 2004; Murthy et al., 2007; Tammiruusu et al., 2007) [132]
Vnitřní prostředí inkluze C. trachomatis
- All ions assayed within the lumenal space of the inclusion approximated the concentrations within the cytoplasm.
- Stimulation of purinergic receptors by addition of extracellular ATP
- Triggered a dynamic Ca2+ response that occurred simultaneously within the cytoplasm and interior of the inclusion.
- The chlamydial inclusion thus appears to be freely permeable to cytoplasmic ions.
- May contribute to the non-fusogenicity of the inclusion with endocytic compartments. [172]
Diferenciace v retikulární tělíska - “reticulate body" (RB) - EB to RB transition
- 8 h p.i., differentiation into RBs is evidenced
- Some EBs containing a condensed nucleoid may still be present
- 12 h p.i., the morphologically typical RBs are observed
- 19 h p.i., multiplication is in full speed.
- EB se mění v metabolically active reticulate body
- Noninfectious intracellular
- Vegetative cell type
- RB is the larger
- Divides by binary fission
- Forms a microcolony referred to as a chlamydial inclusion
- After a period of growth and division
- RB reorganize and condense [25]
- Form new EB [25]
- By 8–12 h reorganization of the EB into RB via numerous morphologically intermediate stages is complete [99]
- 24 to 36 h p.i. the RBs continue to multiply within the inclusion and no EBs are yet detected
- De novo protein expression is required to begin intracellular growth
- Presence of the bacterial histone-like proteins HctA and HctB
- Render the EB transcriptionally incompetent [52]
- Expression of HctA at the start of the cycle would seem to block new transcriptional activity
- Same way this occurs late in the cycle [52]
- Chlamydial histone–DNA interactions are disrupted upon germination by
- A small metabolite in the non-mevalonate pathway (MEP) of isoprenoid biosynthesis
- Thought to be 2-C-methylerythritol 2,4-cyclodiphosphate
- Involved in functional antagonism of HctA [52]
- Chlamydia possess another histone-like protein HctB
- Still unknown what antagonizes its function during the differentiation step [52]
Aktivace chlamydiální ATPázy
- Oligomycin senzitivní
- Mg dependentní
- F-type ATPáza [32]
- Katalyzuje vznik ADP z ATP
- Homologická k mitohcondriální ATPázám
- Redukce ATP produkované mitochondrií [32] - pouze teoretická úvaha
- Pokles přenosu el. v dýchacím řetězic mtch jako důsledek
- Důsledkem může být sekundární porfýrie [32]
- X syntézy hemu, kumulace porfyrinů
- X synt. cytochormu (vč. P-450), peroxidázy, superoxid-dismutáza, hemoglobin, myoglobin, tryptofan pyrrolase,... [32]
- Po lýze infikované buňky uvolnění poryfyrinů do ECT
- V případě infekce jater Chlamydií - možné vyšší koncentrace porfyrinů v hepato-GIT oběhu - sekundární porfyrie [33]
- Zdokumenotván je vzestup tvroby ATP v buňce a v samotné chlamydii [34]
Aktivace dalších genů
- Other genes expressed at 1 h PI included genes that encode proteins involved in
- Translocation of metabolites into the bacterial cell
- ADP/ATP translocase
- Nucleotide phosphate transporter
- Oligopeptide permease
- D-alanine/glycine permease
- Metabolite interconversions
- Malate dehydrogenase
- Nucleoside phosphohydrolase
- methionine aminopeptidase
- Inclusion modification
- Inc-like genes CT228
- CT229
- EEA1-like CT147 [52]
- Unknown functions [52]
- All genes were conserved among chlamydial species [52]
- Genes expressed during primary differentiation serve
- Establishing systems involved in nutrient acquisition
- Modifying the parasitophorous vacuole (inclusion) to prevent its entry into the endocytic pathway leading to lysosomal fusion [52]
Reticular body - RB
- Po primary differentiation
- Larger than EBs (ca. 1 µm)
- Cytoplasm appears granular with diffuse, fibrillar nucleic acids
- Bounded by an inner and outer-membrane
- Resembling other, Gram-negative, eubacteria
- Surface covered with projections and rosettes
- Extend from the bacterial surface through the inclusion membrane
- Proposed TTSS needle structures
- Similar to those found on EBs but at a higher density
- Undergo binary fission
- Throughout the middle part of the developmental cycle [52]
- Certain species RBs tend to be closely associated with the inner face of the inclusion membrane throughout the period of rapid growth
- C. pneumoniae appears to completely fill the interior of the inclusion [52]
- C. caviae tends to grow in an “articulated” form of the inclusion, without a large internal space [52]
- Metabolically active, but osmotically stable
Membrána RB
- Lack the disulfide cross-linked envelope proteins
Modification of the inclusion
- Once internalized, chlamydiae actively modify the properties of the nascent vacuole
- Normal trafficking through the host endocytic pathway
- Dissociating it from late endosomes and lysosomes
- Inability to detect endosomal and lysosomal markers in the Chlamydial inclusion membrane
- Is non-fusogenic with endosomes or lysosomes [52]
- Inclusions intersect a subset of vesicles from the Golgi apparatus containing
- Sphingomyelin
- cholesterol
- Occur early in the developmental cycle
- Necessary for successful replication of Chlamydia within a host cell
- Chlamydial gene expression is required
- Chlamydial proteins that are likely to be important mediators of these properties
- Secreted into the host cell cytoplasm
- Incorporated into the inclusion membrane .
- Chlamydial inclusions
- Trafficked to the perinuclear region of the host cell
- Within 2 h after entry
- Remain in close proximity to the Golgi apparatus
- Begin to fuse with a subset of host vesicles containing sphingomyelin
- Host cell vesicular trafficking
- Moves toward the minus end of microtubules
- Aggregate at the microtubule-organizing center
- Dependent on Chlamydial protein synthesis
- Rab GTPases
- Key regulators of membrane trafficking
- Seen to be recruited to Chlamydial inclusion membrane
- Absence of Rab5, association of Rab4 and Rab11 with the inclusion membrane
- Inclusions are associated with markers or endosomal domains characteristic of late steps in the recycling pathway
- Chlamydial inclusions intersect the cellular autophagic pathway to acquire nutrients
- Using markers for the autophagy pathway
- MAP-LC3
- Calreticulin
- Autophagic vesicles proved to be in close proximity to the inclusion
- Direct fusion between the two vesicles could not be established
- The inclusion of autophagy inhibitors in the growth medium led to aberrant chlamydial growth
- 3-methyl adenine
- Several amino acids [52]
- Inc proteins
- Found in the inclusion membrane
- Interact with host proteins
- Share very limited amino acid identity among themselves
- Share a common bilobular hydrophobicity motif
- Thought to span the inclusion membrane
- IncA , IncD, IncE and IncF, and IncG
- Transcribed within the first 2 h after internalization
- Candidates for chlamydial factors required for the modification of the nascent inclusions [52]
Replication of RB
- By binary fission
- Forming an intracellular microcolony (inclusion)
- By 20 h, the cysteine-rich proteins in the cell walls of the RBs
- Have become linked by disulphide bonds
- Content of RNA is 3–4 times greater thanthat of DNA
- 70S ribosomes have been formed [99]
- Chlamydia lack an identifiable ftsZ ortholog
- Encodes a protein centrally involved in bacterial cell division
- Found in all other sequenced eubacteria
- Presence of a complete set of genes for the synthesis of peptidoglycan
- Numerous studies reported that Chlamydia lacked peptidoglycan
- Single study reported trace amounts in EBs
- Attempts to identify peptidoglycan in RBs were unsuccessful
- Penicillin and other ß-lactams are inhibitory to chlamydial growth
- Probably target the high molecular weight penicillin binding proteins [52]
- RBs may synthesize small amounts of peptidoglycan
- Role in bacterial cell division
- Perhaps by substituting for the lack of FtsZ in the formation of nascent division septa [52]
- Chlamydial MurA ortholog (UDP-N-acetylglucosamine enopyruvyl transferase)
- Catalyzes the first committed step in peptidoglycan biosynthesis
- Is functional in E. coli
- Found to encode a fosfomycin-resistant form of the enzyme
- Immediately preceding cell division - involvement of peptidoglycan synthesis in cell division [52]
- Inclusion expands [99]
- Occupy most of the cytoplasm of the host cell
- RBs take nutrients and energy from the host cells
- Za určitých podmínek RBs do not re-differentiate directly into EBs
- Form non-replicating ‘persistent bodies’
- Maintain a chronic latent infection [14]
- Aberrant bodies (ABs)
- Considered as a hallmark of the persistent Chlamydia infection (Kern et al., 2009)
- Persistence
- Stage of infection where viable but culture-negative
- Nucleic acid-positive organisms reside in the cells (Beatty et al., 1994; Bin et al., 2000).
- Persistent altered life cycle in which the organism exists as an aberrant body can be induced in vitro
- Through indoleamine 2,3-dioxygenase (IDO) activity, which deprives the pathogen of tryptophan [19]
- Obligate intracellular
- Inability culture the organism in the absence of host cells [1]
- Cell-mediated immunity plays a crucial role in the resolution of chlamydial infections [22]
- Human chlamydial species
- Requirements for tyrosine phosphorylation by Src-family kinases
- Not seen in other chlamydial species [8]
- Cannot however synthesize ATP or GTP
- Must rely on the host cell for ATP [25]
- Dle DNA analýzy si nějaké ATP snad produkovat umí [33]
- Chlamydiae may have the capacity to synthesize ATP [99]
- Chlamydiae may scavenge ATP from cytoplasm of the host cell in the early stage of the developmental cycle [99]
- Chlamydia uses the host cellular nutrients for its own metabolism, for example cholesterol (Carabeo et al., 2003)
- C. pneumoniae induce monocyte oxidation of LDL in a dose dependent manner (Kalayoglu, 1999)
- LDL oxidation was enhanced with longer incubation periods and with increasing concentrations of LDL
- Inhibited by the antioxidant alpha-tocopherol (vitamin E)
- vitamin E inhibits C. pneumoniae-induced macrophage LDL oxidation
- To the perinuclear region and the Golgi area
- Dependent on early chlamydial gene expression [100]
- Inhibit the pro-apoptotic signaling molecules
- Bcl-2 family of proteins
- Cytochrome c
- Caspase-3 pathway (Airenne et al., 2002; Fischer et al., 2004)
- The C. pneumoniae infected host cells were human laryngeal carcinoma cell line Hep2 (Fischer et al., 2004).
- Chlamydia-inhibited apoptosis was blocked upstream of the mitochondrial activation of Bax/Bak (Fischer et al., 2004)
- Depend largely on the cell line in question
- C. pneumoniae has also been shown to induce apoptosis in human coronary artery endothelial cells (Schöier et al., 2006).
- In HL cells (Alvesalo et al., 2008)
- Schisandra lignans upregulate heat shock proteins Hsp25, Hsp70 and Hsp72 (Chiu& Ko, 2004; Wu et al., 2004)
- C. pneumoniae downregulates these genes 12 hours post infection
- Time point where Schisandra lignans display antichlamydial activity
- In HL cells (Alvesalo et al., 2008)
- Measured at 24 hours post infection
- Time point where schisandrin B still showed over 20% statistically significant inhibitory activity
- calcium transport
- One of the key components for successful C. pneumoniae entry into the host cell
- calcium as a target for antichlamydial action
- calcium is a very common signaling transmitter in the human body
- Na+ gradient may well be the only energy source for secondary transport.
- Cytochrome d-type terminal oxidases Na+ cycle-dependent ones (adaptace na extracelulární prostředí ?)
- Bacillus halodurans
- E. coli
- P. aeruginosa
- V. cholerae
- H. influenzae
- C. trachomatis
- C. pneumoniae
- Cytochrome d-type oxidases that are not Na+ pumps and microbes do not require Na+ for growth, consider them to be H+ pumps
- B. subtilis
- Synechocystis spp.
- Campylobacter jejuni
- Rickettsia prowazekii
A proto asi nadbytek solení zhoršuje aterosklerozu ?? !!! Chlamydie roste potom rychleji ???
- ATP synthesis, motility, and solute uptake
- Improve its chances for colonization of the host cells and survival in the host organisms
- V. cholerae, the cells appear to respond to alterations in Na+ circulation by modulating the expression of the main virulence regulon
Možná i Chlamydia pneumonia ?
- Pathogens that utilize the Na+ cycle seem to encode a significant share of permeases that belong to Na+-dependent transporter families
- Solute/sodium symporter family (SSS; TC 2.A.21) - Na+-dependent symporters:
- Na+/proline and Na+/pantothenate permeases PutP and PanF from E. coli
- Na+/glucose symporter SglT from V. parahaemolyticus
- Homologous transporters are encoded in:
- P. gingivalis
- C. pneumoniae
- C. difficile
- N. meningitidis
- N. gonorrhoeae
- P. multocida
- H. influenzae
- H. ducreyi
- A. actinomycetemcomitans
- K. pneumoniae
- P. aeruginosa
- S. enterica serovars Typhi and Paratyphi
- V. cholerae
- Y. pestis
- C. trachomatis, C. pneumoniae, and H. influenzae:
- NQR
- Principal respiratory ionic pump
- Pyrimidine nucleotide transhydrogenase
- Cytochrome bd-type terminal oxidase
- Chlamydia pneumonia a její závislost na Na+ cyklu
- Primary Na+ pump, NQR
- Možný terapeutiký cíl (Ag, Li, aj.) / očkování ?
- Specific inhibitors of primary Na+ pumps (korormicin and Ag+)
- Na+-translocating ionophores (monensin)
- H+- (or Na+)-transporting V-type ATPase - common in bacteria
- Na+/alanine symporters (CT409 and CT735 in C. trachomatis, CPn0876 and CPn0536 in C. pneumoniae)
- Na+/branched-chain amino acid symporter (CT554 and CPn0836)
- Uncharacterized transporter of the neurotransmitter:sodium symporter family (CT231 and CPn0290)
- Phosphate permease PitA (CT692 and CPn0680)
- Glutamate transporter GltS (CT401 and CPn0528)
- ADP/ATP translocase (CT065, CT495, CPn0351, and CPn0614)
- Amino acid-polyamine-organocation family (TC 2.A.3) transporters (CT374, CT216, CPn0282 and CPn1031)
- Specificky enzym C. pneumoniae
- Fructose-bisphosphate sequestered by chlamydial outer protein N (Ishida et al, 2014)
- A functional arginine-agmatine bacterial exchange system
- Protects against host nitric oxide production and innate immunity (takes in host arginine) Giles et al, 2009 [153]
- Linked to a putative bacterial membrane transport protein
- Hydroxylate phenylalanine, tyrosine, and tryptophan into tyrosine, dihydroxyphenylalanine, and 5-hydroxytryptophan
Požití Euthyroxu mi výrzaně zhoršilo artralgie
Často bývá přidružená deprese a x nálady
Může tam být nějaká souvislost ?
- Expressed within 24 h of infection
- Hydroxylate host stores of aromatic amino acids
- + during the period of logarithmic bacterial growth [11]
- Maintenance of this gene within a number of Chlamydia
- May have an important role in shaping the metabolism or overall pathogenesis of these bacteria [11]
- All three AroAA-Hs are tetrahydropterin dependent [11]
- Homologs of phenylalanine hydroxylase (PheH)
- Present in about 20% of the bacterial genomes that have been sequenced [11]
- Bacterial PheH differ from mammalian PheH
- Lack the regulatory and tetramerization domains
- But residues key for ligand and metal binding are conserved between eukaryotes and bacteria [11]
- Iron binding [11]
- Appear to be transcriptionally linked to an uncharacterized putative bacterial membrane transport protein [11]
- Changes in the trp operon are key to determining the host niche of some Chlamydia
- Modulation of tryptophan availability
- TyrP gene - tyrosine/tryptophan permease
- Influences C. pneumoniae tissue tropism and pathogenicity
- Vascular strains only contain a single copy
- AroAA-H can convert phenylalanine to tyrosine
- Conversion of tyrosine into 4-hydroxyphenylpyruvate
- Dioxygenase is necessary for 4-hydroxyphenylpyruvate catabolic conversion into homogentisate
- Not present in Chlamydia
- Chlamydial AroAA-H also converts tryptophan into 5-hydroxytryptophan
- Not known to metabolize 5-hydroxytryptophan
- Addition of 5-hydroxytryptophan to C. pneumoniae-infected cells has an inhibitory effect on chlamydial infection [11]
- Binding of the IFN-gamma receptor
- Transcriptionally activates the expression of indoleamine-2,3-dioxygenase
- Degradesl-tryptophan to l-kynurenine
- This cytokine-mediated host cell response deprives intracellular chlamydial RBs of tryptophan
- It ultimately prevents their growth and replicative capabilities !!!
- Treatment of epithelial cells with
- High levels of IFN-gamma
- Completely inhibits growth
- Subinhibitory concentrations
- C.p. ztratila schopnost syntetizovat si tryptofan sama
- Má to vliv na to, ve kterých tkáních lépe přežívá - tropismus [19]
- Encoding functional tryptophan synthase may be a survival factor for intracellular chlamydiae [19]
- Mammalian cells lack the ability to biosynthesize tryptophan [19]
- Respiratory strains of C. pneumoniae
- Possess multiple copies of the tyrP gene
- Encoding a tyrosine-tryptophan permease [39]
- Vascular strains
- Encode only one copy [39]
- Presence of extra tyrP copies correlated with
- Increased mRNA levels
- Higher uptake of the substrate tyrosine in respiratory strains [39]
- Reduced capacity for amino acid transport
- May contribute to a greater tendency of vascular strains to become persistent in vivo [39]
- Chlamydia virulence factor Pgp3
- Neutralizes the antichlamydial activity of human cathelicidin LL-37 (Hou et al, 2015) [153]
- Is maintained by
- Functional components of the electron transport chain (ETC)
- Their own V-ATP synthase (McClarty, 1999; Gerard et al., 2002; Skipp et al., 2005) [154]
- Serine protease
- Degradation
- Several eukaryotic components required for MHC Ag I a II expression
- Regulatory factor X5
- Upstream stimulation factor 1 [7]
- Into the host cell cytoplasm
- Important in both persistent and normal infections
- Inhibited CPAF translocation to the host (HEp-2) cell cytoplasm due to:
- IFN-gamma
- Iron deficiency-induced persistence of C. pneumoniae CWL-029 [39]
- Absence of CPAF protease activity on host cell proteins
- Could also reduce the availability of readily transportable amino acids (including tryptophan)
- Therefore contribute to the maintenance of persistence [39]
- Effector protein
- Secreted from the infection vacuole once effective replication has been established
- Cleave host cell proteins for the chlamydial favour [60]
- Induce Golgi fragmentation [60]
- Vital for C. pneumoniae survival
- Targeting CPAF could be an effective strategy
- Disrupt host cell proinflammatory signaling
- Cleaving a NF-?B family transcription factor p65
- Chlamydial virulence factor
- Limiting host cell sensitivity to proinflammatory stimuli [60]
- CPAF crystal structure
- Complex with a protease inhibitor lactocystin
- Rational design of CPAF inhibitors
- Can be expected to cover only acute chlamydial infection [60]
- Translocation of CPAF to host cell cytoplasm
- Inhibited in persistent infection
- Targeting CPAF may not represent an effective strategy for eradication [60]
- Modify major histocompatibility complex expression [100]
- Able to hydroxylate phenylalanine, tyrosine, and tryptophan
- Cpn1046 is expressed within 24 h of infection
- Allowing C. pneumoniae to hydroxylate host stores of aromatic amino acids during the period of logarithmic bacterial growth (Abromaitis et al, 2009) [153]
- Chlamydial genome contains a cytochrome bd oxidase
- Associated with microaerobic respiration in Coxiella burnettii under low oxygen conditions (Omsland et al., 2009) [154]
- Close homologue to a Bacillus subtilis ermC gene
- Encoding a RNA methyltransferase
- Enzymes belonging to RNA (adenine-N6-)-methyltransferases (EC 2.1.1.48)
- Mediating essential functions in ribosomal methylation and structure
- Cíl k terapii [60]
- Whole genome sequencing revealed metabolic genes of glycolysis and pentose phosphate pathway (PPP) (Stephens et al., 1998; Iliffe-Lee and McClarty, 1999; Kalman et al., 1999; McClarty, 1999) [154]
- C.Pneumoniae expresses an external sphingomyelinase
- Converting host sphingomyelin, to ceramide
- Biophysical properties of the two provide the driving force for autoendocytosis of the bacterium (Penate Medina et al, 2014) [153]
- A protein appendage
- Also essential for the infectivity of Chlamydia spp
- Effector protein injection into the host cell cytosol
- Additional chlamydial proteins are secreted into the host cytosol
- Affect immune recognition
- Intracellular survival (TR)
- Elicits numerous proinflammatory responses by host cells
- Actively modify and impair the host immune system
- Prevents or significantly delays chlamydial recognition and efficient elimination [7]
- Cleaves the p65/RelA subunit of NF-kappa B1 into 40- and 22-kDa fragments
- Preventing NF-kappa B activation during chlamydial infection [7]
- Degradation of TRAF3 [7]
- University of AArhus 2D database [153]
- Sequestering the adaptor molecule Act1 to the chlamydial inclusion membrane
- Intercepts the signaling pathway from IL-17R [7]
- Detectable in the inclusion membrane
- Some are secreted into the cytoplasm
- Including the protease-like activity factor (CPAF)
- In the inclusion membrane
- May affect inclusion development
- Avoidance of lysosomal fusion
- Vacuole trafficking
- Nutrient acquisition
- Condensation
- After 48–72 h (PN) [14]
- 48 h p.i. RBs start to differentiate back into EBs
- Asynchronous
- 67h EBs and intermediate developmental forms are detected
- Some typical RBs are still in the process of binary fission
- 60 and 72 h p.i. - increasing percentage of EBs ready for exocytosis to release for subsequent rounds of infection
- RBs is synchronous until approximately 18–24 h post-infection (TR) [1]
- Dedifferentiation to infectious EBs can first be observed (TR) [1]
- 2-fold increases in abundance in the early stages of RB to EB transition
- Amino acid and cofactor biosynthesis
- Ndk, TrxA, Adk, PyrH, and BirA
- Maintenance of cytoplasmic protein function
- GroEL/ES, DnaK, DksA, GrpE, HtrA, ClpP, ClpB, and Map
- Modification of the bacterial cell surface
- CrpA, OmpA, and OmcB
- Energy metabolism
- Tal and Pyk
- Putative transcriptional regulator
- TctD [51]
- Late gene expression - redifferentiate to EBs - “secondary differentiation”
- Dissociation of dividing RBs from the inclusion membrane as a trigger for secondary differentiation
- Removal of the TTSS system needle apparatus from the inner surface of the inclusion membrane
- Suggested as a critical
- Hypothetical mechanism
- Late-cycle genes encode
- Components of the outer-membrane complex
- OmcA and B
- proteins involved in the condensation of the chromosome
- HctA and B
- Extended the number of genes expressed during this stage of development
- OmcAB, hctAB, ltuB, lcrH
- Formation of the highly disulfide-cross-linked outer-membrane complex
- Thioredoxin disulfide isomerases (CT780 and CT783)
- Membrane thiol proteases (mtpA and mtpB)
- May encode “early” proteins [52]
- 70 late genes - Stage III
- Not expressed until 24 h PI
- OmcAB, hctAB, ltuB, lcrH.1
- CT643 and CT660 in addition to hctAB
- Unknown function and found only in chlamydia [52]
- 36–72 h after the infection
- Variations are seen
- Hundreds of infectious progeny are released from each infected cell [120]
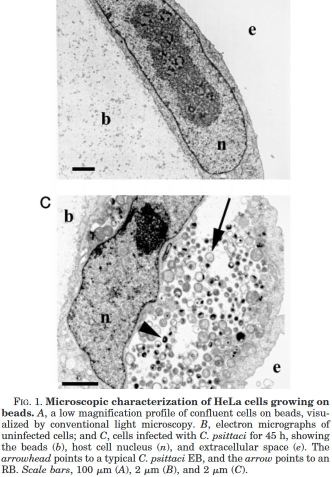
- Na konci růstového cyklu je cytoplazmatický prostor hostitelské buňky téměř vyplněn endozomem – inkluzí
- Počet zralých ET může být až 10 000
- Z napadené buňky se ET dostávají ven buď jejím zánikem či exocytózou [120]
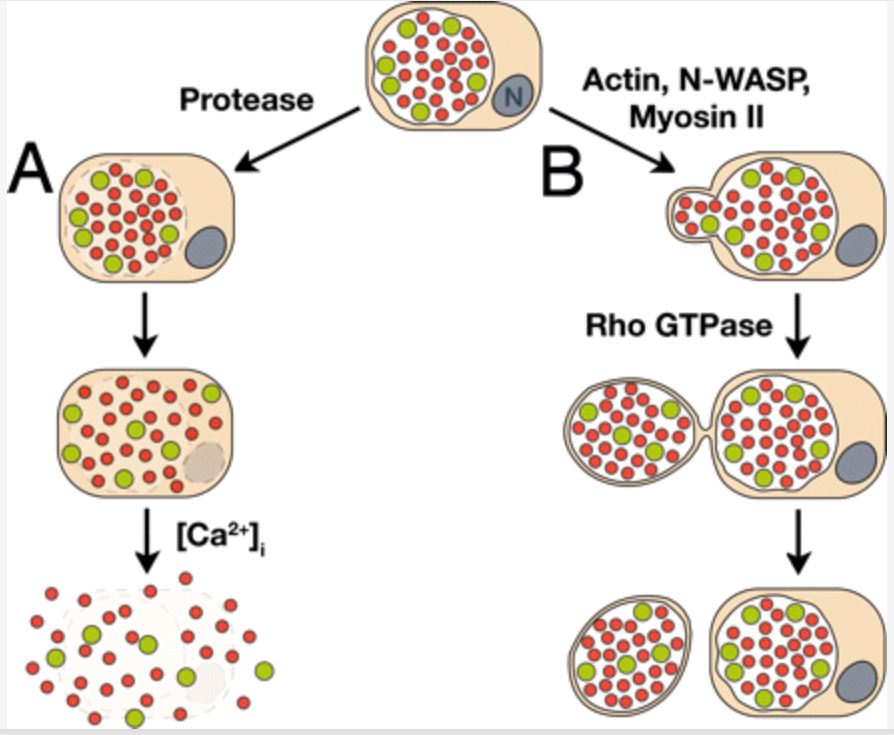
- Packaged release mechanism
- Portion of the chlamydial inclusion was released by a membranous protrusion [168]
- Slow process
- Pinching of the inclusion
- Protrusion out of the cell within a cell membrane compartment
- Shielded from preexisting local immune responses
- Subsequent rupture and release in safer location
- Engulfed by macrophages, thereby facilitating secondary infection and dissemination in the host [168]
- Ultimately detachment from the cell [168]
- Extrusion required
- Actin polymerization
- Alone was sufficient to drive the event [168]
- Neuronal Wiskott–Aldrich syndrome protein
- Myosin II
- Rho GTPase
- The participation of Rho was unique in that it functioned late in extrusion
- Dual nature of release characterized for Chlamydia
- Has not been observed as a strategy for intracellular bacteria [168]
- Extrusions also contained a thin layer of cytoplasm between plasma and inclusion membranes [168]
- Extrusions appeared to project in both lateral and upward directions [168]
- Active reticular body of chlamydiae
- Can spread directly from one host cell to to adjacent cells
- By exploiting lipid raft microdomains in the host cell membrane
- From peripheral blood monocytes to endothelial cells [45]
- Active process
- One would not expect it to take place in the presence of protein synthesis inhibitors
- Cessation of gadolinium leakage in the presence of minocycline in magnetic resonance imaging lesions in MS [45]
- Sequence of membrane permeabilizations
- Inclusion, nucleus and plasma membrane rupture
- Inclusion membrane is disintegrated gradually
- Release of naked EBs into the host cytosol
- Later entry into uninfected host cells
- Inclusions rotate rapidly
- Counterclockwise then clockwise
- Before a volcanic-like rupture to release chlamydial progeny within minutes
- RBs and intermediate forms left at the end of the developmental cycle
- Set free along with the EBs
- Not infectious
- Antigenic stimuli in intact hosts [99]
- Treatment with protease inhibitors abolished inclusion lysis [168]
- Intracellular calcium signaling was shown to be important for plasma membrane breakdown.[168]
- Inhibition of calcium signaling significantly impaired the final step of lysis, plasma membrane rupture
- With no apparent effect on inclusion rupture [168]
- Source of calcium influx was from the extracellular solution and not intracellular stores
- Final step in the lysis pathway of Chlamydia release was calcium-dependent
- Is possible that proteases were also involved [168]
- Chlamydial proteins, including a protease
- Accumulate in the cytoplasm of cells in a temporal manner
- Very late (30 h after infection) gene expression patterns have been reported
- Late-expressed chlamydial protein could induce release [168]
- First infects alveolar macrophages and airway epithelial cells
- Secrete proinflammatory cytokines and chemokines
- Influx of inflammatory cells
- Monocytes, macrophages, and neutrophils
- Could induce human mast cells (MCs)
- To produce cytokines [5]
- Release of IL-8 [49]
- Zvýší plicní permeabilitu pro imunitní buňky
- Rapid transendothelial migration of polymorphonuclear neutrophil granulocytes - PMN [49]
- Required for normal C. pneumoniae propagation [5]
- Epithelial and endothelial cells
- Macrophages / monocytes
- Dendritic cells (DCs) [12]
- Mucosal epithelial cells
- Endothelial
- Smooth muscle cells [48, 99]
- Cardiac muscle cells [48]
- Spleen [99]
- Aorta [99]
- Adipocytes [99]
- Brain cells [99]
- Lymphocytes [99]
- EB infects cells - metabolically inactive form
- 2–6 h after infection developmental cycle begins
- Initial lag phase is seen from 6 to 18 h postinfection
- Next 24–48 h the bacteria grows and replicates - RB form
- Optimal metabolic activity
- Exponential growth from 26 to 46 h [11]
- Replication diminishes
- Individual bacterias - differentiate into the infectious metabolically inactive form of EB
- Different species and serovars at different rates [11]
- Division of RB occurs once every
- 2-3 hours for C. trachomatis [54]
- 6-7 hours for C. pneumoniae [54]
- Most are complete in 40–72 h
- Konec synchronizace výskytu EB a RB
- Host cell lyses
- Infectious progeny are released from the cell [1]
Persistent bodies
Metabolismusm Chl.pn.
Závislost na lidské buňce
Cholesterol
LDL
Trafficking of the EB
Blokáda apoptózy
Indukce apoptozy
Downregulates genes
Chaperone regulator activity
Calcium channel regulatory activity
Energie a protony
Transportní systémy v.s. závislé na H+ gradientu
DhnA-type fructose-1,6-bisphosphate aldolase
ALDOA aldolase A
Dalo by se to využít k diagnostikce ?
Agmatine
AroAA-Hs hydroxyláza
Variations in tryptophan biosynthesis
Je to výhodné pro vyléčení nebo pro vznik perzistence ?
Co se děje, když si dám L-tryptofan? Je to lepší nebo horší?
CAMP cathelicidin antimicrobial peptide
Chlamydial energy generation
Chlamydial protease-like activity factor (CPAF)
C. Pneumoniae protein - Cpn1046
Cytochrome bd oxidase
Dimethyladenosine transferase (coded by the ksgA gene)
Glycolysis enzymes
Sphingomyelinase
T3SS - type 3 secretion system- injectisome
Tail-specific protease
WARS tryptophanyl-tRNA synthetase
Act1
Inc proteins
Cytosol chlamydial proteins
Re-differentiation back into EB = RB to EB transition
EB releas from the infected cell
Exocytosis
Extrusion of the inclusion into neighbouring cells [54]
Transmigration of RB
Host cell lysis
Inclusion lysis
Nucleus lysis
Plasma membrane lysis
Šíření infekce v těle
1. Infekce DC a imunitní reakce
2. Infekce dalších buněk
Developmental cycles - timing